
 科普中國公眾號
科普中國公眾號
 科普中國微博
科普中國微博

 幫助
幫助上世紀八十年代,美國堪薩斯大學的古鳥類學家拉里·馬丁(Larry D. Martin)首次在白堊紀今鳥型類化石中觀察到了齒骨邊緣的一個小小骨茬。根據它所在的位置,馬丁將其命名為前齒骨。
今鳥型類是中生代最為進步的鳥類支系,現生鳥類就是從中演化而來。直至今日,除了在中生代今鳥型類(如燕鳥、義縣鳥、紅山鳥等)中發現了前齒骨,無論是在更原始的類群(比如始祖鳥,孔子鳥或者反鳥類),還是在現生鳥類里,都再也追尋不到前齒骨的蹤影,因此可謂“空前絕后”。
這塊奇特的小骨頭就像是譜寫在演化史上神秘而孤立的一章。當前學術界對于這塊小骨頭的研究,也僅停留在簡要的描述階段,并不了解其來源以及功能。
而馬丁的學生——曾經在堪薩斯大學獲得博士學位、現如今為中科院古脊椎動物與古人類研究所研究員的周忠和,卻從未停止對它的好奇和探究。11月18日,《美國科學院院刊》(PNAS)在線發表了周忠和、Alida Bailleul團隊的研究成果,有望揭開這塊神秘骨頭的面紗。

神秘小骨頭的存在竟是為了獲取食物?
本次研究的材料——熱河生物群的馬氏燕鳥(Yanornis martini)化石,發現于中國遼寧省西部朝陽市的九佛堂組,種名“馬氏”就是為了紀念首次命名前齒骨的古生物學家馬丁。燕鳥的大小與白鴿接近,頭骨很長,嘴里約有30顆牙齒。從燕鳥胃部殘留的魚類骨骸推測,魚類可能是燕鳥的主要食物來源。
此次,當研究者運用骨骼形態觀察、顯微CT以及nano-CT斷層掃描重建、古組織學切片、掃描電子顯微鏡分析等多種手段分析前齒骨時,他們在前齒骨以及齒骨之間發現了特殊的關節軟骨存在的痕跡。
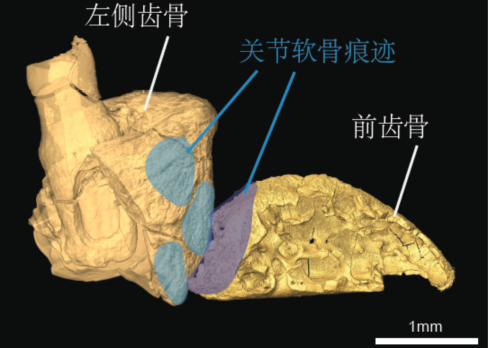
關節軟骨痕跡 (圖片來源:Bailleul et al.,PNAS, 2019)
關節軟骨的存在表明,前齒骨可能通過此關節與齒骨相連,并受下頜神經分支的控制。這個關節可以增加下頜的活動性,而這種增強的活動性顯然與鳥類的取食功能有關。顯然,可動的下頜使得鳥類在獵食的時候擁有了更高度的靈巧性,因此,這一只馬氏燕鳥在捕食它生命中“最后的晚餐”——某條魚的時候,前齒骨無疑發揮了一定的作用。

馬氏燕鳥復原圖 (圖片來源:論文作者提供)
前齒骨的“前身”
基于化石以及已有的現生鳥類的胚胎發育的研究數據,研究人員對前齒骨的來源也進行了探討。
首先,在現生鳥類齒骨骨骼形成的發育過程中,并沒有前齒骨發育的任何線索;其次,研究者推測前齒骨與麥氏軟骨(Meckel’s cartilage, 也稱第一鰓弓軟骨或下頜軟骨)的發育也無直接關系,而應該屬于籽骨(Sesamoid bone)一類的骨骼。
從解剖學上講,籽骨是嵌入肌腱內的骨骼。在人體內,籽骨通常可見于跨關節的肌腱處(如手、膝、足等部位),一般起到保護肌腱和提高其機械性的功能(例如存在于膝關節的髕骨)。而前齒骨就可能起到了相類似生物力學作用。一般情況下,籽骨都生長在頭后骨骼部分,但是也有特殊情況,比如北島垂耳鴉的頭骨內下頜關節部分,就長有三塊籽骨。而前齒骨,可能就是這種長在頭骨的特殊籽骨。
也就是說,前齒骨并非起源于頜骨中任何已有的骨骼,它的出現代表今鳥類演化的一個特有支系中的創新特征。但是鑒于前齒骨的化石很小,保存也更為罕見,這一推測仍有待進一步研究予以驗證。
特殊的“頭骨可動性”
除了關節軟骨,研究者還在前齒骨的組織切片里,發現了角質喙存在的證據。
角質喙主要存在于現生鳥類,保護著齒骨和上頜骨的最前端。馬氏燕鳥的前齒骨很可能曾經被角質喙所包裹。不僅如此,前齒骨的存在和今鳥類嘴部最前端的缺齒性有一定的關聯,在上前頜骨末端與與前齒骨相互咬合的部分,牙齒顯然是缺失的,這部分可能和前齒骨一樣,也被角質喙所包裹,并能夠感知外界的受力激發的信號。
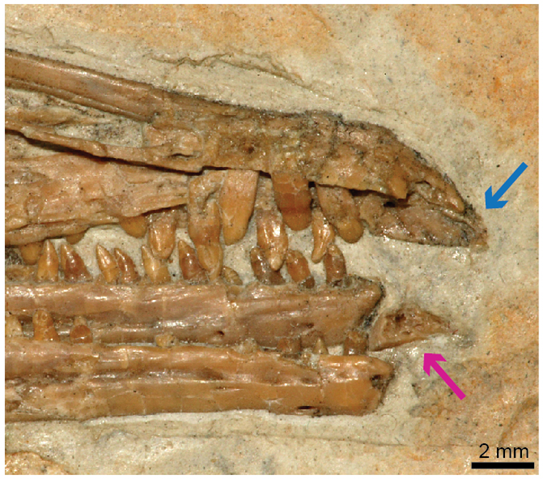
馬氏燕鳥 (IVPP V13358)頭骨前部照片。紅箭頭為前齒骨,藍箭頭是和前齒骨咬合的前上頜骨的缺齒部位。(圖片來源:Bailleul et al.,PNAS, 2019)
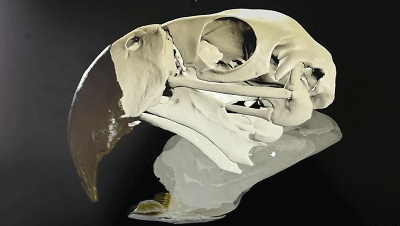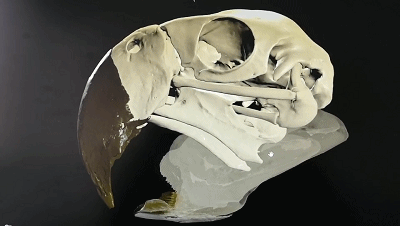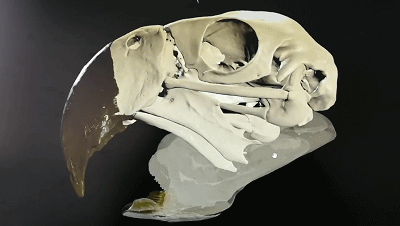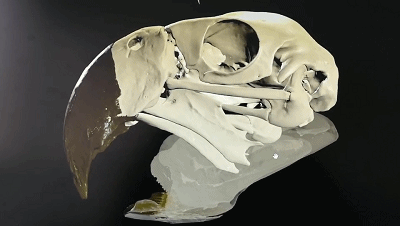
五彩金剛鸚鵡的頭骨可動性3D示意圖
(圖片來源:https://sketchfab.com/3d-models/scarlet-macaw-cranial-kinesis-animation-5a21c8c4c0ac4eb8a98323ed24d68e75)
“頭骨可動性”是主要發生在頭骨內上頜和腦顱之間的運動,在現生鳥類中很普遍,許多鳥類在前頜骨、鼻骨、額骨關聯處可以發生一定程度的相對運動。這一特征可能是鳥類能夠演化至今的重要優勢之一,但是相關證據在白堊紀的古鳥類中卻很少出現。
本次研究證明,馬氏燕鳥上頜骨末端的角質喙、前齒骨、牙齒、齒骨,展現了一種極具特色、但是已經不復存在的、高度靈活的白堊紀古鳥類頭骨可動性(cranial kinesis)。這種特殊的“頭骨可動性”可能僅僅局限在滅絕的今鳥型類化石中,并且從早白堊世一直持續至晚白堊世。遺憾的是,在之后的演化過程中,這種方式逐漸消失,而這一特殊的“頭骨可動性”究竟是如何運作的,成為一道新的科學難題。
不過,本次研究已經顯示出將高精度顯微CT數據與古組織研究結合的學術潛力。也許在未來,類似的演化難題有望被研究者們逐一解答。
參考文獻:
1.Martin L (1987) The beginning of the modern avian radiation. Documents des Laboratoires de Géologie de Lyon 99:9-19.
2.Zhou Z & Martin LD (2011) Distribution of the predentary bone in Mesozoic ornithurine birds. Journal of Systematic Palaeontology 9(1):25-31.
3.Cunningham SJ, et al. (2013) The anatomy of the bill tip of kiwi and associated somatosensory regions of the brain: comparisons with shorebirds. Plos One 8(11):e80036.
4.Bailleul AM, Hall BK, & Horner JR (2012) First Evidence of Dinosaurian Secondary Cartilage in the Post-Hatching Skull of Hypacrosaurus stebingeri (Dinosauria, Ornithischia). Plos One 7(4):e36112.
6. 5. Bailleul AM, Li ZH, O’Connor JK, & Zhou ZH (2019) Origin of the avian predentary and evidence of a unique form of cranial kinesis in Cretaceous ornithuromorphs.PNAS.DOI: 10.1073/pnas.1911820116
7. 維基百科
