
 科普中國(guó)公眾號(hào)
科普中國(guó)公眾號(hào)
 科普中國(guó)微博
科普中國(guó)微博

 幫助
幫助
 一芳田自然科普
一芳田自然科普 一、殺菌作用和抑菌作用1.1 中毒病菌的癥狀病原菌中毒的癥狀主要表現(xiàn)為:菌絲生長(zhǎng)受阻、畸形、扭曲等;孢子不能萌發(fā);各種子實(shí)體、附著孢不能形成;細(xì)胞膨脹、原生質(zhì)瓦解、細(xì)胞壁破壞;病菌長(zhǎng)期處于靜止?fàn)顟B(tài)。1.2 殺菌和抑菌的區(qū)別從中毒癥狀看,殺菌主要表現(xiàn)為孢子不能萌發(fā),而抑菌表現(xiàn)為菌絲生長(zhǎng)受阻(不是死亡),藥劑解除后即可恢復(fù)生長(zhǎng)。從作用機(jī)制看,殺菌主要是影響了生物氧化能的生成,因孢子萌發(fā)需要較多的能量,而抑菌主要是影響了生物合成,菌絲生長(zhǎng)耗能較少。但殺菌和抑菌作用往往不能截然分開(kāi),一個(gè)殺菌劑表現(xiàn)殺菌作用還是抑菌作用,主要和藥劑本身性質(zhì)、藥劑濃度和藥劑作用時(shí)間有關(guān)。從藥劑本身性質(zhì)看,一般重金屬鹽類(lèi)、有機(jī)硫類(lèi)殺菌劑多表現(xiàn)為殺菌作用,而許多內(nèi)吸性殺菌劑,特別是農(nóng)用抗菌素則常表現(xiàn)為抑菌作用。從藥劑濃度看,低濃度通常表現(xiàn)為抑菌作用,而高濃度時(shí)則表現(xiàn)為殺菌作用。從藥劑作用時(shí)間看,作用時(shí)間短常表現(xiàn)為抑菌作用,延長(zhǎng)作用時(shí)間,則表現(xiàn)為殺菌作用。二、殺菌劑的主要作用部位和機(jī)制從生物化學(xué)角度講,殺菌劑的作用機(jī)制主要是影響了病原菌的生物氧化;或是影響了病原菌的生物合成。2.1 影響生物氧化的殺菌劑2.1.1 生物氧化的生化背景病原菌的生命過(guò)程需要能量,尤其是孢子萌發(fā),更需要較多的能量供應(yīng),這些能量來(lái)自碳水化合物、脂肪和蛋白質(zhì)的氧化生成的ATP。其中碳水化合物的氧化尤為重要,糖的氧化主要有糖酵解通路、有氧氧化通路和磷酸戊糖支路。糖酵解通路,是不需氧的呼吸過(guò)程,是葡糖或葡糖-1-磷酸變成丙酮酸的過(guò)程,而在缺氧情況下,糖酵解則包括丙酮酸進(jìn)一步還原成乳酸(在哺乳動(dòng)物中)或經(jīng)乙醛還原成乙醇(如在酵母菌中),糖酵解是在細(xì)胞質(zhì)中進(jìn)行的。有氧氧化通路:其前面一大段和糖酵解相同,只是到磷酸甘油醛時(shí),才在輔酶Ⅰ的作用下脫氫,經(jīng)二磷酸甘油酸形成丙酮酸乃至乙酰輔酶A,然后進(jìn)入三羧酸循環(huán),轉(zhuǎn)入電子傳遞鏈及末端氧化。其中,三羧酸循環(huán)是在線(xiàn)粒體的基質(zhì)中進(jìn)行的,而電子傳遞則在線(xiàn)粒體“嵴”(即內(nèi)膜)上進(jìn)行的。磷酸戊糖支路:從6-磷酸葡糖經(jīng)6-磷酸葡糖酸到戊糖的旁路,因此又稱(chēng)磷酸戊糖之路,磷酸戊糖之路也是在細(xì)胞質(zhì)中進(jìn)行的。脂肪酸氧化時(shí),先在細(xì)胞質(zhì)中活化,生成酯酰輔酶A,活化了的脂肪酸進(jìn)入線(xiàn)粒體進(jìn)行β氧化,生成的乙酰輔酶A進(jìn)入三羧酸循環(huán)。三羧酸循環(huán)Krebs cycle是所有有機(jī)物質(zhì)初步代謝產(chǎn)物的共同氧化途徑,最終產(chǎn)生CO2和水,具體過(guò)程如圖1 三羧酸循環(huán)示意圖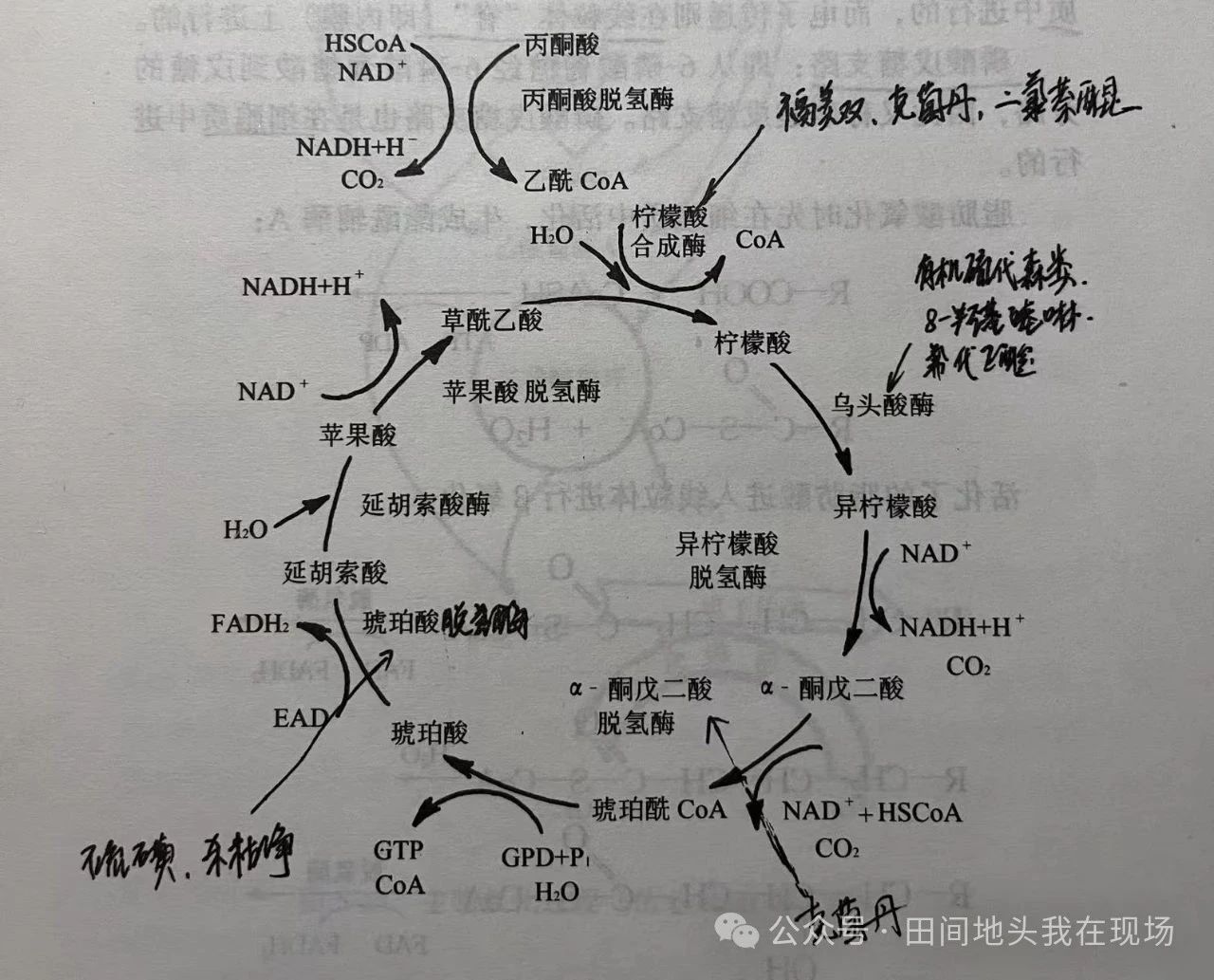 圖1 三羧酸循環(huán)示意圖呼吸鏈電子傳遞是在線(xiàn)粒體的脊上進(jìn)行的,目前,可將線(xiàn)粒體的這一亞單位劃分成4種復(fù)合體,吸氧化作用產(chǎn)生的能量不是直接被利用的,而是暫時(shí)被貯存在高能化合物如三磷酸腺苷ATP中,然后再由高能化合物釋放出來(lái)(如當(dāng)ATP-ADP時(shí))。呼吸氧化產(chǎn)生的能量進(jìn)入高能化合物一般涉及兩個(gè)過(guò)程,一個(gè)是氧化過(guò)程,另一個(gè)是磷酸化過(guò)程,即能量作為磷的高能鍵貯存起來(lái),這兩個(gè)耦合的過(guò)程即為氧化磷酸化。2.1.2 酞酰亞胺類(lèi)殺菌劑作用機(jī)制酞酰亞胺類(lèi)主要有克菌丹captan、滅菌丹folpet和敵菌丹captafol,是廣譜殺菌劑,主要作用機(jī)制有三種,一是影響丙酮酸的脫羧作用,使之不能進(jìn)入三羧酸循環(huán);二是抑制a-酮戊二酸脫氫酶的活性,阻斷三羧酸循環(huán);三是作用于含-SH的酶或輔酶。影響丙酮酸的脫羧作用,使之不能進(jìn)入三羧酸循環(huán)。主要表現(xiàn)為:棉鈴紅腐病菌用克菌丹處理后,發(fā)現(xiàn)其細(xì)胞內(nèi)丙酮酸大量積累,而很少有乙酰輔酶A生成,實(shí)質(zhì)是克菌丹改變了丙酮酸脫氫酶系中一種輔酶硫胺素TPP。硫胺素在丙酮酸脫羧過(guò)程中的作用是轉(zhuǎn)移乙酰基。輔酶硫胺素TPP的關(guān)鍵結(jié)構(gòu)是噻唑環(huán)中氮和硫原子之間的碳原子上的氫很容易離解,使該碳原子形成反應(yīng)性很強(qiáng)的負(fù)碳離子,因而可親核攻擊丙酮酸的羰基原子形成加成物。輔酶硫氨酸TPP的噻唑環(huán)上的氮帶正電子,可作為電子受體使脫羧容易進(jìn)行,脫羧后產(chǎn)生羥乙基TPP,TPP經(jīng)反應(yīng)后噻唑環(huán)上的氮不再帶正電荷,沒(méi)有接受電子的能力,也就失去了轉(zhuǎn)移乙酰基的功能。抑制a-酮戊二酸脫氫酶系的活性,阻斷三羧酸循環(huán)。三羧酸循環(huán)中,從a-酮戊二酸到琥珀酰輔酶A需要a-酮戊二酸脫氫酶系催化,而這一酶系的一種輔酶也是硫胺素TPP,因此和上述丙酮酸脫氫酶系的情形相同,克菌丹也作用于TPP,從而阻斷了三羧酸循環(huán)。作用于含-SH的酶或輔酶。不但破壞了輔酸A,而且生成的硫光氣還能抑制酶或輔酶的活性,因?yàn)榱蚬鈿庖子诤偷鞍踪|(zhì)中的-SH,-OH,-NH2等基團(tuán)反應(yīng),因此,克菌丹等酞酰胺類(lèi)殺菌劑是多作用點(diǎn)的殺菌劑。2.1.3 硫代氨基甲酸酯類(lèi)殺菌劑的作用機(jī)制硫代氨基甲酸酯類(lèi)殺菌劑主要包括“代森”系列和“福美”系列,其作用機(jī)制主要包括破壞輔酶A和抑制以銅、鐵等為輔基的酶的活性。破壞輔酶A,代森鋅和福美雙都有類(lèi)似反應(yīng)。輔酶A被瓦解后直接影響了脂肪酸的β氧化,丙酮酸脫氫酶系、a-酮戊二酸脫氫酶系的活性受到抑制,因?yàn)檫@些酶系中必須要有輔酶A的參與。抑制以銅、鐵等為輻基的酶的活性。硫代氨基甲酸酯類(lèi)殺菌劑可和鐵、銅等形成螯合物可使酶失去活性。如在三羧酸循環(huán)中,檸檬酸經(jīng)順烏頭酸到異檸檬酸必須要有烏頭酸酶的參與,而烏頭酸酶的輔基含有高鐵,代森類(lèi)、福美類(lèi)殺菌劑和鐵形成螯合物使烏頭酸酶失活,三羧酸循環(huán)中斷。2.1.4 取代苯類(lèi)殺菌劑的作用機(jī)制取代苯類(lèi)殺菌劑以百菌清chlorothalonil為代表,還有diclroan和dichlone,其主要作用機(jī)制在于和含-SH的酶反應(yīng),抑制了含-SH基團(tuán)酶的活性,特別是磷酸甘油醛脫氫酶的活性。磷酸甘油醛脫氫酶催化糖酵解途徑中從3-磷酸甘油醛到1,3-二磷酸甘油酸的反應(yīng)。其催化機(jī)理是磷酸甘油醛脫氫酶活性位置上半胱氨酸殘基的-SH基是親核基團(tuán),它與醛基作用形成中間產(chǎn)物,可將羥基上的氫轉(zhuǎn)移至與酶緊密結(jié)合的NAD+上,從而產(chǎn)生NADH和高能硫酯中間產(chǎn)物。NADH從酶上解離,另外的NAD+與酶活性中心結(jié)合,磷酸攻擊硫酯鍵從而形成1,3-二磷酸甘油。百菌清和該酶的-SH結(jié)合,抑制其活性,中斷糖酵解,從而影響ATP的生成。此外,也有觀點(diǎn)認(rèn)為百菌清也和含-SH的谷胱甘肽反應(yīng),破壞了谷胱甘肽。谷胱甘肽在菌體內(nèi)對(duì)外源物的解讀反應(yīng)中有主要作用,抗百菌清菌株中的谷胱甘肽含量遠(yuǎn)遠(yuǎn)高于敏感菌株,這也是百菌清作用于谷胱甘肽的一個(gè)證據(jù)。2.1.5 羧酰苯胺類(lèi)殺菌劑作用機(jī)制羧酰苯胺類(lèi),以氧硫雜環(huán)二烯為主,還有噻吩、噻唑、呋喃、吡唑、苯基等衍生物,代表作品有萎銹靈carboxin、氧化萎銹靈oxycarboxin、鄰酰胺mebenil、氟酰胺flutolanil、furametper、triflumazid等。這些殺菌劑的主要作用部位是線(xiàn)粒體呼吸電子傳遞鏈中從琥珀酸輔酶Q之間的氧化還原體系,即復(fù)合體Ⅱ。復(fù)合體Ⅱ是由黃酶Ⅱ(FAD)為輔酶的黃素蛋白、非血紅素鐵硫蛋白和其他結(jié)合蛋白組成的。底物(琥珀酸)脫出的2個(gè)H傳遞給FAD-FADH2,而三價(jià)鐵離子將FADH2氧化成FAD,放出2個(gè)H,同時(shí)三價(jià)鐵離子還原為二價(jià)鐵離子,這2個(gè)H又被輔酶Q接受,稱(chēng)為輔酶QH2,萎銹靈和復(fù)合體Ⅱ的活性中心—非血紅素鐵硫蛋白結(jié)合,從而阻斷了電子向輔酶Q的傳遞。氟酰胺等殺菌劑也作用于復(fù)合體Ⅱ,但這些殺菌劑的結(jié)合部位既不是黃素蛋白,也不是非血紅素鐵硫蛋白這兩個(gè)主要的亞單位,而是結(jié)合于一種固膜蛋白,嵌入鐵硫蛋白和輔酶Q之間,阻止電子傳遞。2.1.6 甲氧丙烯酸酯類(lèi)殺菌劑的作用機(jī)制甲氧丙烯酸酯類(lèi)殺菌劑是以Strobilurin A為先導(dǎo)化合物,開(kāi)發(fā)出的新型殺菌劑,代表品種有ICIA5504和BAS490F,其作用機(jī)理是抑制了病原菌線(xiàn)粒體呼吸電子傳遞鏈中電子的傳遞,其作用部位是復(fù)合體Ⅲ(即細(xì)胞色素b和細(xì)胞色素C1復(fù)合體)。通過(guò)和復(fù)合體Ⅲ中的活性部位結(jié)合而抑制了線(xiàn)粒體的電子傳遞。此外,抗菌素抗霉素A antimycinA和殺菌劑殺枯凈phenazine,5-氧吩嗪也是作用于復(fù)合體Ⅲ。2.1.7 敵克松的作用機(jī)制敵克松dexon為重氮磺酸鹽類(lèi)殺菌劑,主要用于防治煙草黑脛病、小麥腥黑穗病、白菜軟腐病和水稻爛秧。敵克松作用于復(fù)合體Ⅰ,阻斷了輔酶Ⅰ(NAD)和黃酶Ⅰ(FMN)之間的電子傳遞。磷酸核黃素FMN中具有異咯嗪環(huán)結(jié)構(gòu),敵克松可能和這種異咯嗪環(huán)組成了一個(gè)穩(wěn)定的復(fù)合物,從而使磷酸核黃素FMN失去傳遞電子的功能。2.1.8 氟啶胺的作用機(jī)制氟啶胺fluazinam是一種強(qiáng)有力的解偶聯(lián)劑,破壞氧化磷酸化,推測(cè)是分子中的氨基基團(tuán)的質(zhì)子化和質(zhì)子化作用引起的。五氯硝基苯terrachlor也是解偶聯(lián)劑。2.2 影響生物合成的殺菌劑2.2.1 有機(jī)磷殺菌劑的作用機(jī)制有機(jī)磷殺菌劑以異稻瘟凈和克瘟散為代表,主要用于防治水稻稻瘟病,20世紀(jì)80年以后人們傾向于認(rèn)為這類(lèi)殺菌劑主要是抑制了卵磷脂的合成而破壞了細(xì)胞質(zhì)膜的結(jié)構(gòu),卵磷脂(磷脂酰膽堿)是細(xì)胞質(zhì)膜最重要的組分,卵磷脂的合成必需要有磷脂酰甲基轉(zhuǎn)移酶的參與,異稻瘟凈的作用機(jī)制主要是抑制磷脂酰乙醇氨甲基轉(zhuǎn)移酶的活性,阻斷了卵磷脂合成。2.2.2 嘧啶胺類(lèi)殺菌劑作用機(jī)制早期開(kāi)發(fā)的嘧啶胺類(lèi)殺菌劑有甲菌啶dimethirimol和乙菌啶ethirimol,主要用于防治瓜類(lèi)和谷物白粉病,乙菌啶主要是非競(jìng)爭(zhēng)性地抑制了腺(嘌呤核)苷脫氨酶的活性而影響了某些堿基和核酸的合成。腺苷脫氨酶是在某一組織中的核酸分解后的堿基可以被另一組重新利用起來(lái)合成新的核酸。其他嘧啶苯胺類(lèi)產(chǎn)品如嘧菌胺mepanipyrim,pyrimethanil和cyprodinil,對(duì)灰葡萄孢引起的多種病害,特別是灰霉病有特效,而且與二甲酰亞胺類(lèi)殺菌劑無(wú)交互抗性。目前對(duì)嘧菌胺等殺菌劑的作用機(jī)制主要有兩種解釋?zhuān)环N是抑制細(xì)胞壁降解酶的分泌,二是干擾甲硫氨酸的生物合成,其抑制了甲硫氨酸生物合成途徑中次末端——β胱硫醚裂解酶的活性,從而抑制了甲硫氨酸的合成,但詳細(xì)抑制機(jī)理還不清楚。2.2.3 苯基酰胺類(lèi)殺菌劑作用機(jī)制苯基酰胺類(lèi)殺菌劑至少包括4類(lèi):酰基丙氨酸類(lèi)、丁內(nèi)酯類(lèi)、硫代丁內(nèi)酯類(lèi)和噁唑烷酮類(lèi),其中以酰基丙氨酸類(lèi)(以甲霜靈為代表)、噁唑烷酮類(lèi)(以噁霜靈為代表)最重要,這類(lèi)殺菌劑廣泛應(yīng)用于藻菌綱病害(如霜霉病)的防治。苯基酰胺類(lèi)殺菌劑的作用機(jī)理一般認(rèn)為是抑制了病原菌中核酸的生物合成,主要是RNA的合成。細(xì)胞各類(lèi)RNA,包括參與翻譯過(guò)程的mRNA、rRNA和tRNA,以及具有特殊功能的小RNA,都是以DNA為模板,在RNA聚合酶的催化下合成的,真核生物的RNA聚合酶有好多種,分子量大約在50萬(wàn)/u,通常由4-6種亞基組成,并含有鋅離子。利用抑制劑a-鵝膏蕈堿的抑制作用可將其分為3類(lèi),對(duì)抑制劑不敏感的RNA聚合酶A(或Ⅰ),可被低濃度抑制劑抑制的RNA B(或Ⅱ),只被高濃度抑制劑抑制的RNA C(或Ⅲ),有觀點(diǎn)認(rèn)為,甲霜靈、噁霜靈主要是抑制了對(duì)a-鵝膏蕈堿不敏感的RNA聚合酶A,從而阻礙了rRNA前體的轉(zhuǎn)錄,具體的抑制機(jī)理不清楚。2.2.4 農(nóng)用抗菌素的作用機(jī)制多氧霉素D的作用機(jī)制多氧霉素D(polyoxin D)主要干擾真菌幾丁質(zhì)的合成,幾丁質(zhì)是構(gòu)成某些真菌細(xì)胞壁的主要組分,其生物合成過(guò)程如下:葡萄糖—6-磷酸葡萄糖胺—N-乙酰葡萄糖胺-6-磷酸—N-乙酰葡萄糖胺-1-磷酸—脲苷二磷酸N-乙酰葡萄糖胺—(幾丁質(zhì)合成酶)幾丁質(zhì)。由于多氧霉素D和尿苷二磷酸N-乙酰葡萄糖胺分子結(jié)構(gòu)有一定相似性,多氧霉素D和底物競(jìng)爭(zhēng)性地結(jié)合幾丁質(zhì)合成酶,從而抑制了該酶的活性,破壞了真菌細(xì)胞壁的結(jié)構(gòu)。井崗霉素的作用機(jī)制井崗霉素,又稱(chēng)有效霉素,主要是有效霉素A,是水溶性?xún)?nèi)吸殺菌劑,而且很容易在紋枯病菌絲中傳導(dǎo),引起菌絲異常分枝,從而抑制其生長(zhǎng)。作用機(jī)制主要是抑制核酸和蛋白質(zhì)合成,井岡霉素對(duì)菌體中海藻糖酶有拮抗性抑制作用,因此提出其作用機(jī)制是基于阻斷了海藻糖分解而切斷了葡萄糖供應(yīng),海藻糖把一個(gè)海藻糖分子分解成2個(gè)葡萄糖。春雷霉素和滅瘟素的作用機(jī)制主要是抑制了病原菌的蛋白質(zhì)合成,蛋白質(zhì)合成的場(chǎng)所是核糖體rRNA,蛋白質(zhì)合成的大致過(guò)程如下:氨基酸活化,消耗一個(gè)ATP,氨基酸和tRNA結(jié)合;mRNA結(jié)合到rRNA的30S小亞基上;攜帶氨基酸的兩個(gè)tRNA結(jié)合到30S小亞基上,大亞基也與之結(jié)合形成穩(wěn)定的70S復(fù)合體;在轉(zhuǎn)肽酶作用下,將P位點(diǎn)的氨基酸轉(zhuǎn)移到A位點(diǎn)的帶氨基酸的tRNA上,與之形成肽鍵;核糖體沿mRNA相對(duì)移動(dòng)一個(gè)密碼的距離,供P位點(diǎn)上脫去氨基的tRNA從A位點(diǎn)移到P位點(diǎn),空出A位點(diǎn),接受下一個(gè)帶氨基酸的tRNA,從而使肽鏈延長(zhǎng);到一定長(zhǎng)度時(shí),停止合成,多肽脫下,mRNA和rRNA分開(kāi),大小亞基分開(kāi);滅瘟素是和rRNA的大亞基上某一單一位點(diǎn)結(jié)合,從而影響了氨酰-tRNA和其應(yīng)有的位點(diǎn)結(jié)合,阻止了肽鏈延長(zhǎng);春雷霉素也和30S小亞基結(jié)合,從而阻止30S小亞基與氨酰-tRNA以及mRNA形成復(fù)合物,蛋白質(zhì)合成無(wú)法起始。2.2.5 麥角甾醇合成抑制劑作用機(jī)制麥角甾醇etgosterol是植物細(xì)胞膜的重要組分,其合成受阻將間接的影響細(xì)胞膜的通透性功能,此外,麥角甾醇還是甾類(lèi)激素的前體,在無(wú)性、有性生殖過(guò)程中起重要作用。目前至少有8類(lèi),幾十個(gè)殺菌劑品種的作用機(jī)制是抑制麥角甾醇合成。其中,三唑類(lèi)、嘧啶類(lèi)、咪唑類(lèi)和哌嗪類(lèi)抑制了24-甲撐-24,25-二氫羊毛甾醇到4,4-二甲基fecosterol C-14脫甲基的反應(yīng);嗎啉類(lèi)主要是抑制了異構(gòu)酶活性;嗎啉/哌啶類(lèi)殺菌劑在菌體生理pH條件下可質(zhì)子化,模擬了碳陽(yáng)離子中間態(tài)和這兩種酶強(qiáng)烈結(jié)合,從而使酶失活。2.2.6 苯并咪唑類(lèi)殺菌劑作用機(jī)制苯并咪唑類(lèi)殺菌劑,苯來(lái)特、多菌靈和甲基硫菌靈等殺菌劑主要影響菌體內(nèi)微管的形成而影響了細(xì)胞分裂。在植物體內(nèi),苯來(lái)特和硫菌靈都轉(zhuǎn)換成多菌靈起作用,主要作用機(jī)制是由于多菌靈和微管蛋白的β亞單位相結(jié)合,阻止了微管的組裝,從而破壞了紡錘體的形成,影響了細(xì)胞分裂。2.2.7 三環(huán)唑和豐谷隆的作用機(jī)制殺菌劑三環(huán)唑tricyclelazole、咯嗪酮pyroquilon、稻瘟醇、四氯苯酞、chlobenthiazone等都主要用于防治水稻稻瘟病,起作用機(jī)制主要用于影響了黑素的生物合成。菌類(lèi)黑素Velanin是一類(lèi)酚類(lèi)化合物,黑素在病原菌的致病性中起主要作用,稻梨孢和豆刺盤(pán)孢與葫蘆科刺盤(pán)孢對(duì)植物侵染前先形成一種附著孢的結(jié)構(gòu),然后該結(jié)構(gòu)穿透寄主表皮細(xì)胞壁而產(chǎn)生侵染,在附著孢穿透表皮之前不久,這些附著孢的壁黑化,如果附著孢的壁不黑化,則不會(huì)穿透表皮或其他屏障,因此三環(huán)唑等又稱(chēng)為抗穿透劑。也有觀點(diǎn)認(rèn)為,三環(huán)唑可能是抑制了多聚乙酰的代謝,造成多聚乙酰的積累,而多聚乙酰對(duì)稻瘟病菌來(lái)說(shuō)是有毒物質(zhì),亦或抑制多聚乙酰代謝和阻斷黑素合成相輔相成。2.2.8 苯基吡咯殺菌劑的作用機(jī)制以天然抗生素硝吡咯菌素pyrrolnitrin為先導(dǎo)化合物開(kāi)發(fā)出一類(lèi)新型殺菌劑,拌種咯和fludioxonil是起代表,有觀點(diǎn)認(rèn)為,蛋白激酶PK-Ⅲ是苯基吡咯殺菌劑的初始靶標(biāo),蛋白激酶PK-Ⅲ調(diào)節(jié)體內(nèi)甘油的合成。拌種咯和蛋白激酶PK-Ⅲ結(jié)合,抑制了它的活性,使活化的調(diào)節(jié)蛋白不失活(不被磷酸化),從而導(dǎo)致甘油合成失控,細(xì)胞內(nèi)滲透壓加大,細(xì)胞發(fā)生腫脹而死亡。2.3 防御素激活劑的作用機(jī)制這一類(lèi)殺菌劑對(duì)病原菌在離體條件下并無(wú)殺菌作用,但可以誘導(dǎo)病原菌的寄主植物產(chǎn)生某些防御作用物質(zhì),即植物防御素,從而防止病害的蔓延。乙磷鋁aliette可以有效的防治藻菌綱病害,但在離體條件下并不表現(xiàn)明顯的殺菌作用,主要是通過(guò)干擾植物多元酚代謝,刺激寄主植物產(chǎn)生酚類(lèi)和倍半萜烯類(lèi)植物防御素,從而抑制了病害的擴(kuò)展。噻菌靈probenazole對(duì)稻瘟病和白葉枯病有良好的防治效果,噻菌靈和其在稻株中的主要代謝產(chǎn)物糖精(鄰磺酰苯甲酰亞胺)和水楊酸,可以促使有毒酯類(lèi)物質(zhì)的產(chǎn)生并提高過(guò)氧化物酶的水平,阻止病菌擴(kuò)展。水楊酸類(lèi)似物用于白粉病、葉瘟病和煙草霜霉病的防治,作用機(jī)制同樣是激活了寄主植物的防御系統(tǒng)。
圖1 三羧酸循環(huán)示意圖呼吸鏈電子傳遞是在線(xiàn)粒體的脊上進(jìn)行的,目前,可將線(xiàn)粒體的這一亞單位劃分成4種復(fù)合體,吸氧化作用產(chǎn)生的能量不是直接被利用的,而是暫時(shí)被貯存在高能化合物如三磷酸腺苷ATP中,然后再由高能化合物釋放出來(lái)(如當(dāng)ATP-ADP時(shí))。呼吸氧化產(chǎn)生的能量進(jìn)入高能化合物一般涉及兩個(gè)過(guò)程,一個(gè)是氧化過(guò)程,另一個(gè)是磷酸化過(guò)程,即能量作為磷的高能鍵貯存起來(lái),這兩個(gè)耦合的過(guò)程即為氧化磷酸化。2.1.2 酞酰亞胺類(lèi)殺菌劑作用機(jī)制酞酰亞胺類(lèi)主要有克菌丹captan、滅菌丹folpet和敵菌丹captafol,是廣譜殺菌劑,主要作用機(jī)制有三種,一是影響丙酮酸的脫羧作用,使之不能進(jìn)入三羧酸循環(huán);二是抑制a-酮戊二酸脫氫酶的活性,阻斷三羧酸循環(huán);三是作用于含-SH的酶或輔酶。影響丙酮酸的脫羧作用,使之不能進(jìn)入三羧酸循環(huán)。主要表現(xiàn)為:棉鈴紅腐病菌用克菌丹處理后,發(fā)現(xiàn)其細(xì)胞內(nèi)丙酮酸大量積累,而很少有乙酰輔酶A生成,實(shí)質(zhì)是克菌丹改變了丙酮酸脫氫酶系中一種輔酶硫胺素TPP。硫胺素在丙酮酸脫羧過(guò)程中的作用是轉(zhuǎn)移乙酰基。輔酶硫胺素TPP的關(guān)鍵結(jié)構(gòu)是噻唑環(huán)中氮和硫原子之間的碳原子上的氫很容易離解,使該碳原子形成反應(yīng)性很強(qiáng)的負(fù)碳離子,因而可親核攻擊丙酮酸的羰基原子形成加成物。輔酶硫氨酸TPP的噻唑環(huán)上的氮帶正電子,可作為電子受體使脫羧容易進(jìn)行,脫羧后產(chǎn)生羥乙基TPP,TPP經(jīng)反應(yīng)后噻唑環(huán)上的氮不再帶正電荷,沒(méi)有接受電子的能力,也就失去了轉(zhuǎn)移乙酰基的功能。抑制a-酮戊二酸脫氫酶系的活性,阻斷三羧酸循環(huán)。三羧酸循環(huán)中,從a-酮戊二酸到琥珀酰輔酶A需要a-酮戊二酸脫氫酶系催化,而這一酶系的一種輔酶也是硫胺素TPP,因此和上述丙酮酸脫氫酶系的情形相同,克菌丹也作用于TPP,從而阻斷了三羧酸循環(huán)。作用于含-SH的酶或輔酶。不但破壞了輔酸A,而且生成的硫光氣還能抑制酶或輔酶的活性,因?yàn)榱蚬鈿庖子诤偷鞍踪|(zhì)中的-SH,-OH,-NH2等基團(tuán)反應(yīng),因此,克菌丹等酞酰胺類(lèi)殺菌劑是多作用點(diǎn)的殺菌劑。2.1.3 硫代氨基甲酸酯類(lèi)殺菌劑的作用機(jī)制硫代氨基甲酸酯類(lèi)殺菌劑主要包括“代森”系列和“福美”系列,其作用機(jī)制主要包括破壞輔酶A和抑制以銅、鐵等為輔基的酶的活性。破壞輔酶A,代森鋅和福美雙都有類(lèi)似反應(yīng)。輔酶A被瓦解后直接影響了脂肪酸的β氧化,丙酮酸脫氫酶系、a-酮戊二酸脫氫酶系的活性受到抑制,因?yàn)檫@些酶系中必須要有輔酶A的參與。抑制以銅、鐵等為輻基的酶的活性。硫代氨基甲酸酯類(lèi)殺菌劑可和鐵、銅等形成螯合物可使酶失去活性。如在三羧酸循環(huán)中,檸檬酸經(jīng)順烏頭酸到異檸檬酸必須要有烏頭酸酶的參與,而烏頭酸酶的輔基含有高鐵,代森類(lèi)、福美類(lèi)殺菌劑和鐵形成螯合物使烏頭酸酶失活,三羧酸循環(huán)中斷。2.1.4 取代苯類(lèi)殺菌劑的作用機(jī)制取代苯類(lèi)殺菌劑以百菌清chlorothalonil為代表,還有diclroan和dichlone,其主要作用機(jī)制在于和含-SH的酶反應(yīng),抑制了含-SH基團(tuán)酶的活性,特別是磷酸甘油醛脫氫酶的活性。磷酸甘油醛脫氫酶催化糖酵解途徑中從3-磷酸甘油醛到1,3-二磷酸甘油酸的反應(yīng)。其催化機(jī)理是磷酸甘油醛脫氫酶活性位置上半胱氨酸殘基的-SH基是親核基團(tuán),它與醛基作用形成中間產(chǎn)物,可將羥基上的氫轉(zhuǎn)移至與酶緊密結(jié)合的NAD+上,從而產(chǎn)生NADH和高能硫酯中間產(chǎn)物。NADH從酶上解離,另外的NAD+與酶活性中心結(jié)合,磷酸攻擊硫酯鍵從而形成1,3-二磷酸甘油。百菌清和該酶的-SH結(jié)合,抑制其活性,中斷糖酵解,從而影響ATP的生成。此外,也有觀點(diǎn)認(rèn)為百菌清也和含-SH的谷胱甘肽反應(yīng),破壞了谷胱甘肽。谷胱甘肽在菌體內(nèi)對(duì)外源物的解讀反應(yīng)中有主要作用,抗百菌清菌株中的谷胱甘肽含量遠(yuǎn)遠(yuǎn)高于敏感菌株,這也是百菌清作用于谷胱甘肽的一個(gè)證據(jù)。2.1.5 羧酰苯胺類(lèi)殺菌劑作用機(jī)制羧酰苯胺類(lèi),以氧硫雜環(huán)二烯為主,還有噻吩、噻唑、呋喃、吡唑、苯基等衍生物,代表作品有萎銹靈carboxin、氧化萎銹靈oxycarboxin、鄰酰胺mebenil、氟酰胺flutolanil、furametper、triflumazid等。這些殺菌劑的主要作用部位是線(xiàn)粒體呼吸電子傳遞鏈中從琥珀酸輔酶Q之間的氧化還原體系,即復(fù)合體Ⅱ。復(fù)合體Ⅱ是由黃酶Ⅱ(FAD)為輔酶的黃素蛋白、非血紅素鐵硫蛋白和其他結(jié)合蛋白組成的。底物(琥珀酸)脫出的2個(gè)H傳遞給FAD-FADH2,而三價(jià)鐵離子將FADH2氧化成FAD,放出2個(gè)H,同時(shí)三價(jià)鐵離子還原為二價(jià)鐵離子,這2個(gè)H又被輔酶Q接受,稱(chēng)為輔酶QH2,萎銹靈和復(fù)合體Ⅱ的活性中心—非血紅素鐵硫蛋白結(jié)合,從而阻斷了電子向輔酶Q的傳遞。氟酰胺等殺菌劑也作用于復(fù)合體Ⅱ,但這些殺菌劑的結(jié)合部位既不是黃素蛋白,也不是非血紅素鐵硫蛋白這兩個(gè)主要的亞單位,而是結(jié)合于一種固膜蛋白,嵌入鐵硫蛋白和輔酶Q之間,阻止電子傳遞。2.1.6 甲氧丙烯酸酯類(lèi)殺菌劑的作用機(jī)制甲氧丙烯酸酯類(lèi)殺菌劑是以Strobilurin A為先導(dǎo)化合物,開(kāi)發(fā)出的新型殺菌劑,代表品種有ICIA5504和BAS490F,其作用機(jī)理是抑制了病原菌線(xiàn)粒體呼吸電子傳遞鏈中電子的傳遞,其作用部位是復(fù)合體Ⅲ(即細(xì)胞色素b和細(xì)胞色素C1復(fù)合體)。通過(guò)和復(fù)合體Ⅲ中的活性部位結(jié)合而抑制了線(xiàn)粒體的電子傳遞。此外,抗菌素抗霉素A antimycinA和殺菌劑殺枯凈phenazine,5-氧吩嗪也是作用于復(fù)合體Ⅲ。2.1.7 敵克松的作用機(jī)制敵克松dexon為重氮磺酸鹽類(lèi)殺菌劑,主要用于防治煙草黑脛病、小麥腥黑穗病、白菜軟腐病和水稻爛秧。敵克松作用于復(fù)合體Ⅰ,阻斷了輔酶Ⅰ(NAD)和黃酶Ⅰ(FMN)之間的電子傳遞。磷酸核黃素FMN中具有異咯嗪環(huán)結(jié)構(gòu),敵克松可能和這種異咯嗪環(huán)組成了一個(gè)穩(wěn)定的復(fù)合物,從而使磷酸核黃素FMN失去傳遞電子的功能。2.1.8 氟啶胺的作用機(jī)制氟啶胺fluazinam是一種強(qiáng)有力的解偶聯(lián)劑,破壞氧化磷酸化,推測(cè)是分子中的氨基基團(tuán)的質(zhì)子化和質(zhì)子化作用引起的。五氯硝基苯terrachlor也是解偶聯(lián)劑。2.2 影響生物合成的殺菌劑2.2.1 有機(jī)磷殺菌劑的作用機(jī)制有機(jī)磷殺菌劑以異稻瘟凈和克瘟散為代表,主要用于防治水稻稻瘟病,20世紀(jì)80年以后人們傾向于認(rèn)為這類(lèi)殺菌劑主要是抑制了卵磷脂的合成而破壞了細(xì)胞質(zhì)膜的結(jié)構(gòu),卵磷脂(磷脂酰膽堿)是細(xì)胞質(zhì)膜最重要的組分,卵磷脂的合成必需要有磷脂酰甲基轉(zhuǎn)移酶的參與,異稻瘟凈的作用機(jī)制主要是抑制磷脂酰乙醇氨甲基轉(zhuǎn)移酶的活性,阻斷了卵磷脂合成。2.2.2 嘧啶胺類(lèi)殺菌劑作用機(jī)制早期開(kāi)發(fā)的嘧啶胺類(lèi)殺菌劑有甲菌啶dimethirimol和乙菌啶ethirimol,主要用于防治瓜類(lèi)和谷物白粉病,乙菌啶主要是非競(jìng)爭(zhēng)性地抑制了腺(嘌呤核)苷脫氨酶的活性而影響了某些堿基和核酸的合成。腺苷脫氨酶是在某一組織中的核酸分解后的堿基可以被另一組重新利用起來(lái)合成新的核酸。其他嘧啶苯胺類(lèi)產(chǎn)品如嘧菌胺mepanipyrim,pyrimethanil和cyprodinil,對(duì)灰葡萄孢引起的多種病害,特別是灰霉病有特效,而且與二甲酰亞胺類(lèi)殺菌劑無(wú)交互抗性。目前對(duì)嘧菌胺等殺菌劑的作用機(jī)制主要有兩種解釋?zhuān)环N是抑制細(xì)胞壁降解酶的分泌,二是干擾甲硫氨酸的生物合成,其抑制了甲硫氨酸生物合成途徑中次末端——β胱硫醚裂解酶的活性,從而抑制了甲硫氨酸的合成,但詳細(xì)抑制機(jī)理還不清楚。2.2.3 苯基酰胺類(lèi)殺菌劑作用機(jī)制苯基酰胺類(lèi)殺菌劑至少包括4類(lèi):酰基丙氨酸類(lèi)、丁內(nèi)酯類(lèi)、硫代丁內(nèi)酯類(lèi)和噁唑烷酮類(lèi),其中以酰基丙氨酸類(lèi)(以甲霜靈為代表)、噁唑烷酮類(lèi)(以噁霜靈為代表)最重要,這類(lèi)殺菌劑廣泛應(yīng)用于藻菌綱病害(如霜霉病)的防治。苯基酰胺類(lèi)殺菌劑的作用機(jī)理一般認(rèn)為是抑制了病原菌中核酸的生物合成,主要是RNA的合成。細(xì)胞各類(lèi)RNA,包括參與翻譯過(guò)程的mRNA、rRNA和tRNA,以及具有特殊功能的小RNA,都是以DNA為模板,在RNA聚合酶的催化下合成的,真核生物的RNA聚合酶有好多種,分子量大約在50萬(wàn)/u,通常由4-6種亞基組成,并含有鋅離子。利用抑制劑a-鵝膏蕈堿的抑制作用可將其分為3類(lèi),對(duì)抑制劑不敏感的RNA聚合酶A(或Ⅰ),可被低濃度抑制劑抑制的RNA B(或Ⅱ),只被高濃度抑制劑抑制的RNA C(或Ⅲ),有觀點(diǎn)認(rèn)為,甲霜靈、噁霜靈主要是抑制了對(duì)a-鵝膏蕈堿不敏感的RNA聚合酶A,從而阻礙了rRNA前體的轉(zhuǎn)錄,具體的抑制機(jī)理不清楚。2.2.4 農(nóng)用抗菌素的作用機(jī)制多氧霉素D的作用機(jī)制多氧霉素D(polyoxin D)主要干擾真菌幾丁質(zhì)的合成,幾丁質(zhì)是構(gòu)成某些真菌細(xì)胞壁的主要組分,其生物合成過(guò)程如下:葡萄糖—6-磷酸葡萄糖胺—N-乙酰葡萄糖胺-6-磷酸—N-乙酰葡萄糖胺-1-磷酸—脲苷二磷酸N-乙酰葡萄糖胺—(幾丁質(zhì)合成酶)幾丁質(zhì)。由于多氧霉素D和尿苷二磷酸N-乙酰葡萄糖胺分子結(jié)構(gòu)有一定相似性,多氧霉素D和底物競(jìng)爭(zhēng)性地結(jié)合幾丁質(zhì)合成酶,從而抑制了該酶的活性,破壞了真菌細(xì)胞壁的結(jié)構(gòu)。井崗霉素的作用機(jī)制井崗霉素,又稱(chēng)有效霉素,主要是有效霉素A,是水溶性?xún)?nèi)吸殺菌劑,而且很容易在紋枯病菌絲中傳導(dǎo),引起菌絲異常分枝,從而抑制其生長(zhǎng)。作用機(jī)制主要是抑制核酸和蛋白質(zhì)合成,井岡霉素對(duì)菌體中海藻糖酶有拮抗性抑制作用,因此提出其作用機(jī)制是基于阻斷了海藻糖分解而切斷了葡萄糖供應(yīng),海藻糖把一個(gè)海藻糖分子分解成2個(gè)葡萄糖。春雷霉素和滅瘟素的作用機(jī)制主要是抑制了病原菌的蛋白質(zhì)合成,蛋白質(zhì)合成的場(chǎng)所是核糖體rRNA,蛋白質(zhì)合成的大致過(guò)程如下:氨基酸活化,消耗一個(gè)ATP,氨基酸和tRNA結(jié)合;mRNA結(jié)合到rRNA的30S小亞基上;攜帶氨基酸的兩個(gè)tRNA結(jié)合到30S小亞基上,大亞基也與之結(jié)合形成穩(wěn)定的70S復(fù)合體;在轉(zhuǎn)肽酶作用下,將P位點(diǎn)的氨基酸轉(zhuǎn)移到A位點(diǎn)的帶氨基酸的tRNA上,與之形成肽鍵;核糖體沿mRNA相對(duì)移動(dòng)一個(gè)密碼的距離,供P位點(diǎn)上脫去氨基的tRNA從A位點(diǎn)移到P位點(diǎn),空出A位點(diǎn),接受下一個(gè)帶氨基酸的tRNA,從而使肽鏈延長(zhǎng);到一定長(zhǎng)度時(shí),停止合成,多肽脫下,mRNA和rRNA分開(kāi),大小亞基分開(kāi);滅瘟素是和rRNA的大亞基上某一單一位點(diǎn)結(jié)合,從而影響了氨酰-tRNA和其應(yīng)有的位點(diǎn)結(jié)合,阻止了肽鏈延長(zhǎng);春雷霉素也和30S小亞基結(jié)合,從而阻止30S小亞基與氨酰-tRNA以及mRNA形成復(fù)合物,蛋白質(zhì)合成無(wú)法起始。2.2.5 麥角甾醇合成抑制劑作用機(jī)制麥角甾醇etgosterol是植物細(xì)胞膜的重要組分,其合成受阻將間接的影響細(xì)胞膜的通透性功能,此外,麥角甾醇還是甾類(lèi)激素的前體,在無(wú)性、有性生殖過(guò)程中起重要作用。目前至少有8類(lèi),幾十個(gè)殺菌劑品種的作用機(jī)制是抑制麥角甾醇合成。其中,三唑類(lèi)、嘧啶類(lèi)、咪唑類(lèi)和哌嗪類(lèi)抑制了24-甲撐-24,25-二氫羊毛甾醇到4,4-二甲基fecosterol C-14脫甲基的反應(yīng);嗎啉類(lèi)主要是抑制了異構(gòu)酶活性;嗎啉/哌啶類(lèi)殺菌劑在菌體生理pH條件下可質(zhì)子化,模擬了碳陽(yáng)離子中間態(tài)和這兩種酶強(qiáng)烈結(jié)合,從而使酶失活。2.2.6 苯并咪唑類(lèi)殺菌劑作用機(jī)制苯并咪唑類(lèi)殺菌劑,苯來(lái)特、多菌靈和甲基硫菌靈等殺菌劑主要影響菌體內(nèi)微管的形成而影響了細(xì)胞分裂。在植物體內(nèi),苯來(lái)特和硫菌靈都轉(zhuǎn)換成多菌靈起作用,主要作用機(jī)制是由于多菌靈和微管蛋白的β亞單位相結(jié)合,阻止了微管的組裝,從而破壞了紡錘體的形成,影響了細(xì)胞分裂。2.2.7 三環(huán)唑和豐谷隆的作用機(jī)制殺菌劑三環(huán)唑tricyclelazole、咯嗪酮pyroquilon、稻瘟醇、四氯苯酞、chlobenthiazone等都主要用于防治水稻稻瘟病,起作用機(jī)制主要用于影響了黑素的生物合成。菌類(lèi)黑素Velanin是一類(lèi)酚類(lèi)化合物,黑素在病原菌的致病性中起主要作用,稻梨孢和豆刺盤(pán)孢與葫蘆科刺盤(pán)孢對(duì)植物侵染前先形成一種附著孢的結(jié)構(gòu),然后該結(jié)構(gòu)穿透寄主表皮細(xì)胞壁而產(chǎn)生侵染,在附著孢穿透表皮之前不久,這些附著孢的壁黑化,如果附著孢的壁不黑化,則不會(huì)穿透表皮或其他屏障,因此三環(huán)唑等又稱(chēng)為抗穿透劑。也有觀點(diǎn)認(rèn)為,三環(huán)唑可能是抑制了多聚乙酰的代謝,造成多聚乙酰的積累,而多聚乙酰對(duì)稻瘟病菌來(lái)說(shuō)是有毒物質(zhì),亦或抑制多聚乙酰代謝和阻斷黑素合成相輔相成。2.2.8 苯基吡咯殺菌劑的作用機(jī)制以天然抗生素硝吡咯菌素pyrrolnitrin為先導(dǎo)化合物開(kāi)發(fā)出一類(lèi)新型殺菌劑,拌種咯和fludioxonil是起代表,有觀點(diǎn)認(rèn)為,蛋白激酶PK-Ⅲ是苯基吡咯殺菌劑的初始靶標(biāo),蛋白激酶PK-Ⅲ調(diào)節(jié)體內(nèi)甘油的合成。拌種咯和蛋白激酶PK-Ⅲ結(jié)合,抑制了它的活性,使活化的調(diào)節(jié)蛋白不失活(不被磷酸化),從而導(dǎo)致甘油合成失控,細(xì)胞內(nèi)滲透壓加大,細(xì)胞發(fā)生腫脹而死亡。2.3 防御素激活劑的作用機(jī)制這一類(lèi)殺菌劑對(duì)病原菌在離體條件下并無(wú)殺菌作用,但可以誘導(dǎo)病原菌的寄主植物產(chǎn)生某些防御作用物質(zhì),即植物防御素,從而防止病害的蔓延。乙磷鋁aliette可以有效的防治藻菌綱病害,但在離體條件下并不表現(xiàn)明顯的殺菌作用,主要是通過(guò)干擾植物多元酚代謝,刺激寄主植物產(chǎn)生酚類(lèi)和倍半萜烯類(lèi)植物防御素,從而抑制了病害的擴(kuò)展。噻菌靈probenazole對(duì)稻瘟病和白葉枯病有良好的防治效果,噻菌靈和其在稻株中的主要代謝產(chǎn)物糖精(鄰磺酰苯甲酰亞胺)和水楊酸,可以促使有毒酯類(lèi)物質(zhì)的產(chǎn)生并提高過(guò)氧化物酶的水平,阻止病菌擴(kuò)展。水楊酸類(lèi)似物用于白粉病、葉瘟病和煙草霜霉病的防治,作用機(jī)制同樣是激活了寄主植物的防御系統(tǒng)。
來(lái)源: 公眾號(hào):田間地頭我在現(xiàn)場(chǎng)
