
 科普中國(guó)公眾號(hào)
科普中國(guó)公眾號(hào)
 科普中國(guó)微博
科普中國(guó)微博

 幫助
幫助
 中國(guó)科普博覽
中國(guó)科普博覽 
出品:科普中國(guó)
作者:呂澤龍(中國(guó)科學(xué)院動(dòng)物研究所)
監(jiān)制:中國(guó)科普博覽
提起鳥(niǎo)類,你腦海中會(huì)浮現(xiàn)怎樣的畫面?是展翅高飛的雄鷹,還是城市公園中隨處可見(jiàn)的麻雀?還有些了解古生物的人會(huì)立刻聯(lián)想到——恐龍。沒(méi)錯(cuò),基于化石材料和系統(tǒng)分類學(xué),鳥(niǎo)類正是恐龍總目(Dinosauria)、蜥臀目、獸腳亞目的分支。并且,一些獸腳亞目恐龍也和鳥(niǎo)一樣長(zhǎng)有羽毛。
不過(guò),與現(xiàn)代鳥(niǎo)類的羽毛相比,這些恐龍的羽毛形態(tài)更加多樣:有些類似頭發(fā)、雞鴨的絨羽,有些則是片狀對(duì)稱或不對(duì)稱羽毛。例如,科學(xué)家們基于雷神翼龍(Tupandactylus)等翼龍的化石研究,發(fā)現(xiàn)翼龍?bào)w表就覆有絲絨狀的羽毛。
這些羽毛究竟從何而來(lái)?在鳥(niǎo)類、非鳥(niǎo)類恐龍以及翼龍身上,羽毛可能扮演著多重角色——或許是飛行的工具,也許是求偶時(shí)的華麗裝飾,又或者是作為偽裝的外衣。本篇文章,讓我們一同深入探索其中的奧秘。
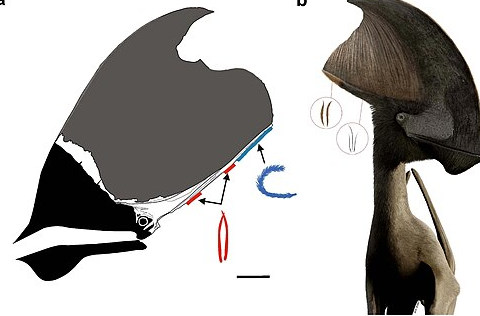
雷神翼龍及其原始羽毛
(圖片來(lái)源:維基百科)

獸腳亞目恐龍羽王龍(Yutyrannus)的尾部化石,保留有羽毛證據(jù)
(圖片來(lái)源:維基百科)
羽毛形態(tài)的演變之路
其實(shí),羽毛最初的形態(tài)并非我們所熟知的那樣,而是有點(diǎn)類似于絲絨或者頭發(fā)。不過(guò)它們的組成成分與人類頭發(fā)并不一樣,人類頭發(fā)主要成分是阿爾法角蛋白,而它們的成分里面主要是貝塔角蛋白。
早期的羽毛是從皮膚表面延伸出來(lái)的絲狀結(jié)構(gòu),呈中空?qǐng)A柱形且沒(méi)有分叉,可能用于偽裝或者裝飾,也有人認(rèn)為這樣的功能可以維持體溫。之后,這些羽毛的結(jié)構(gòu)逐漸改變——分叉且呈現(xiàn)絲狀。再之后出現(xiàn)了羽軸,又形成對(duì)稱的片狀羽毛。

具有片狀羽毛的赫氏近鳥(niǎo)龍標(biāo)本
(圖片來(lái)源:作者拍攝于國(guó)家自然博物館)
圓柱形且中空的絲狀結(jié)構(gòu)羽毛在天宇龍、鸚鵡嘴龍、古林達(dá)奔龍身上發(fā)現(xiàn)過(guò),這些恐龍并非蜥臀目的獸腳類恐龍,而是屬于鳥(niǎo)臀目恐龍。這表明,這些原始的羽毛可能起源于鳥(niǎo)臀目恐龍、蜥臀目恐龍和翼龍的最近共同祖先,只不過(guò)在后續(xù)演化道路上,有些成員失去了羽毛,比如體型龐大的三角龍和真蜥腳類恐龍。而另一些類群則進(jìn)一步強(qiáng)化了羽毛的發(fā)育,如上文提到的雷神翼龍,就有絲狀不分叉羽毛與絲狀分叉羽毛兩種。

原始中華龍鳥(niǎo)化石模型,有原始的絲狀羽毛
(圖片來(lái)源:作者拍攝于國(guó)家自然博物館)

天宇龍復(fù)原圖
(圖片來(lái)源dinosaurpictures.org)
在獸腳類恐龍的廓羽盜龍類(Pennaraptora)分支中(包括竊蛋龍類、鳥(niǎo)翼類、馳龍科和傷齒龍科等),演化出了對(duì)稱片狀羽毛,并帶有“羽軸”——羽毛的“脊梁骨”。雖說(shuō)這些羽毛的空氣動(dòng)力學(xué)性能仍然有限,但是可以在求偶時(shí)用來(lái)展示。廓羽盜龍類的胡氏耀龍(Epidexipteryx hui)就是典型的例子,學(xué)界認(rèn)為其尾巴上明顯的修長(zhǎng)尾羽可能就是裝飾,用以吸引異性。

珍珠雞羽毛,紅色圈內(nèi)較粗的“分割線”為羽軸
(圖片來(lái)源:維基百科)

胡氏耀龍標(biāo)本,有尾巴羽毛
(圖片來(lái)源:維基百科)
當(dāng)片狀羽毛演化出不對(duì)稱結(jié)構(gòu)時(shí),往往預(yù)示著飛行能力的出現(xiàn)。仔細(xì)觀察這些片狀羽毛的兩側(cè),會(huì)發(fā)現(xiàn)羽毛的側(cè)面有很多“毛”,這些類似枝條的“毛”稱之為羽枝。羽枝又是由帶著溝槽的羽小枝構(gòu)成的,羽小枝盤根錯(cuò)節(jié)地排列,依靠它們的羽小鉤互相“咬”在一起形成溝槽連鎖機(jī)制。羽小枝排在一起的同時(shí),這些“鎖”不能在受到外力時(shí)散開(kāi),為此,羽小枝上還帶有結(jié)節(jié)結(jié)構(gòu)來(lái)增強(qiáng)固定。為什么要固定上呢?因?yàn)橹挥泄潭ㄆ饋?lái)才可以讓用于飛行的羽毛足夠“強(qiáng)韌”,抗撕裂。
抗撕裂的飛行羽毛并非所有恐龍的共同特征。在廓羽盜龍類中,傷齒龍科(Troodontidae)和馳龍科(Dromaeosauridae)這些近鳥(niǎo)類非鳥(niǎo)恐龍雖然擁有片狀羽毛,但它們羽毛片中間的羽軸相對(duì)纖細(xì),而且羽小枝排列很松散,沒(méi)有強(qiáng)韌的連鎖結(jié)構(gòu)。
反鳥(niǎo)類屬于鳥(niǎo)類但是并非現(xiàn)代鳥(niǎo)類,它們則擁有變厚的羽軸這一新特征,但是羽小枝之間排列依舊不夠緊密,相鄰的羽小枝之間存在空隙,這就導(dǎo)致它們飛行空氣動(dòng)力學(xué)效率低。相比之下,這些活到今天的恐龍后裔——現(xiàn)存的鳥(niǎo)類,其羽毛能抗撕裂,羽小枝緊密排列在一起,飛行效率顯著較高。

反鳥(niǎo)類的代表類群之一——渤海鳥(niǎo)類
(圖片來(lái)源:作者拍攝于國(guó)家自然博物館)

顧氏小盜龍化石
(圖片來(lái)源:作者拍攝于國(guó)家自然博物館)
近期研究中,科學(xué)家發(fā)現(xiàn)始祖鳥(niǎo)具有三級(jí)飛羽,而很多類似鳥(niǎo)的非鳥(niǎo)類恐龍則沒(méi)有這個(gè)結(jié)構(gòu)。當(dāng)羽小枝排列較為松散時(shí),就會(huì)形成鳥(niǎo)類和部分非鳥(niǎo)類恐龍擁有的絨羽。鳥(niǎo)類是廓羽盜龍類下面鳥(niǎo)翼類(Avialae)的分支,從晚侏羅世到晚白堊世期間,其骨骼結(jié)構(gòu)經(jīng)歷了顯著變化,物種數(shù)目也有所增加。鳥(niǎo)翼類中的真鳥(niǎo)類分支包括現(xiàn)代鳥(niǎo)類,其胸骨等結(jié)構(gòu)和反鳥(niǎo)類存在差異。
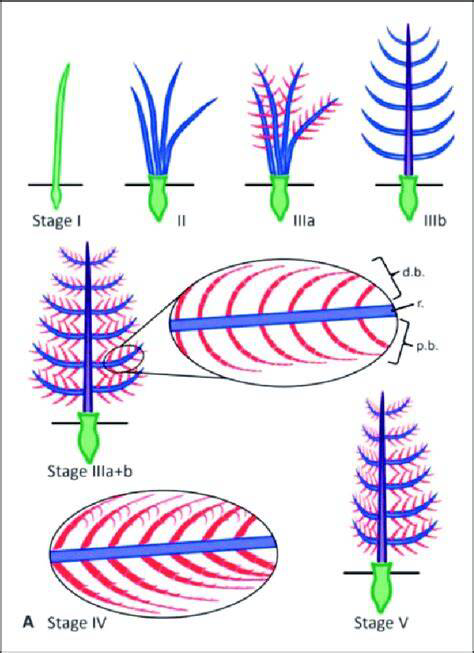
從絲狀纖維(Stage Ⅰ)到分叉(II),再到羽軸和羽小枝形成(IIIa和IIIb),最后羽小枝排列起來(lái)(Stage IV),不對(duì)稱飛羽形成(Stage V)
(圖片來(lái)源:參考文獻(xiàn)[15])
羽毛是如何發(fā)育形成的?
為了找出羽毛發(fā)育形成的奧秘,科學(xué)家們用馴化紅原雞(也就是家雞)的胚胎進(jìn)行了實(shí)驗(yàn),嘗試讓雞全身長(zhǎng)出絲狀的原始羽毛。
如何誘導(dǎo)原始羽毛的生長(zhǎng)呢?關(guān)鍵在于SHH信號(hào)通路,該通路能影響鳥(niǎo)類體表羽毛和裸區(qū)鱗片的發(fā)育。2023年就有科學(xué)家通過(guò)促進(jìn)這一通路,成功使雞腿部原本光滑的鱗片轉(zhuǎn)變?yōu)橛鹈?/p>
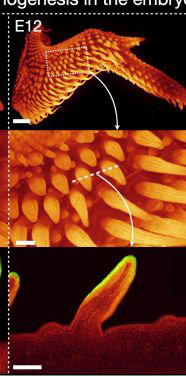
雞胚胎12天的原始羽毛
(圖片來(lái)源:參考文獻(xiàn)[3])

雞爪的鱗片轉(zhuǎn)變?yōu)橛鹈珜?shí)驗(yàn)
(圖片來(lái)源:參考文獻(xiàn)[17])
考慮到雞胚胎在發(fā)育的過(guò)程中,羽毛原基會(huì)發(fā)育成相應(yīng)的羽毛,因此研究人員最初選擇在胚胎發(fā)育第9天(羽毛原基尚未長(zhǎng)成),開(kāi)始用藥物抑制這一基因信號(hào)通路。然而,實(shí)驗(yàn)并未達(dá)到預(yù)期效果——雖然藥物在早期階段顯示出抑制作用,但從雞胚胎第14天起,雞羽毛逐漸發(fā)育為結(jié)構(gòu)復(fù)雜的絨羽和片狀羽毛等。
這一結(jié)果表明,從龍到鳥(niǎo)類的羽毛性狀結(jié)構(gòu)變得復(fù)雜,依賴的是復(fù)雜的基因調(diào)控網(wǎng)絡(luò)的共同作用,調(diào)控網(wǎng)絡(luò)也可以在環(huán)境干擾下保證羽毛發(fā)育。當(dāng)然,在實(shí)驗(yàn)中那些被加入藥物抑制基因信號(hào)通路的雞胚胎,在孵化成為小雞之后,比起未加入藥物的對(duì)照組小雞,身上具有更多的“裸區(qū)”沒(méi)有羽毛。雖說(shuō)這些小雞在成長(zhǎng)的過(guò)程中,身上的“裸區(qū)”也長(zhǎng)出了結(jié)構(gòu)復(fù)雜的羽毛。
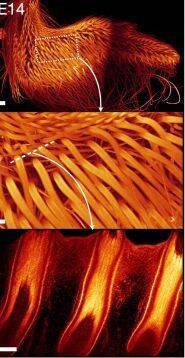
14天的雞羽毛
(圖片來(lái)源:參考文獻(xiàn)[3])
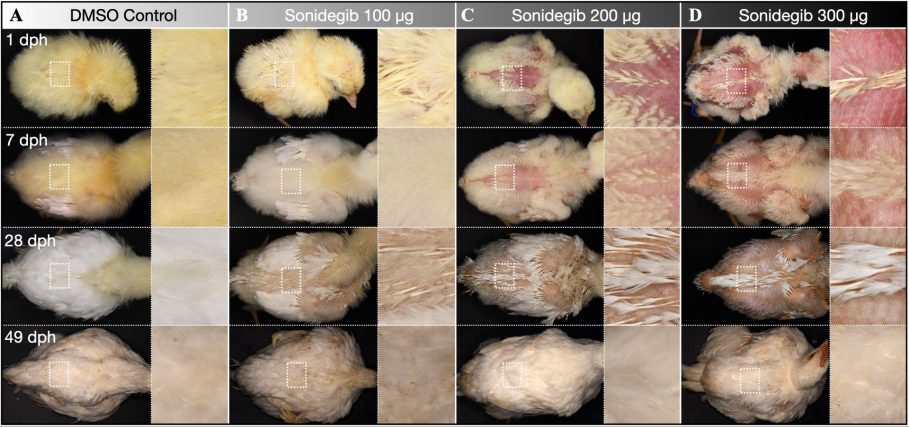
胚胎分別加入不同量抑制藥物的小雞的羽毛生長(zhǎng)情況(從左到右依次是對(duì)照組,100微克,200微克和300微克),后面三個(gè)在孵化之初,體表有明顯較多裸區(qū)
(圖片來(lái)源:參考文獻(xiàn)[3])
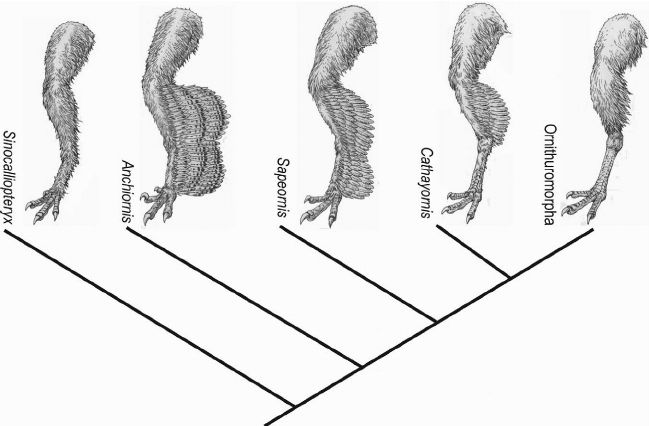
從左到右依次是并非鳥(niǎo)翼類的中國(guó)龍鳥(niǎo),鳥(niǎo)翼類原始的近鳥(niǎo)龍,鳥(niǎo)翼類的會(huì)鳥(niǎo),反鳥(niǎo)類的華夏鳥(niǎo)和現(xiàn)代鳥(niǎo)類所屬分支,可以看出鳥(niǎo)翼類腿上的毛減少了
(圖片來(lái)源:參考文獻(xiàn)[14])
當(dāng)鳥(niǎo)類演化出羽毛時(shí),其體表的鱗片也并非全都消失了。基于對(duì)鳥(niǎo)翼類恐龍(包括鳥(niǎo)類)腿部羽毛的分析發(fā)現(xiàn),原始的鳥(niǎo)翼類恐龍?jiān)诔B(niǎo)類演化過(guò)程中,退化了腿部羽毛,重新發(fā)育出鱗片。這一演化特征在現(xiàn)代家雞的足部鱗片中得到了典型體現(xiàn)。
羽毛的特殊功能
在功能上,科學(xué)家們還發(fā)現(xiàn)了不同鳥(niǎo)類和恐龍羽毛的特別用處。除了我們熟悉的藍(lán)孔雀、紅原雞等鳥(niǎo)類的性展示功能,鳥(niǎo)身上還有纖羽(hair feather)。這類羽毛可以作為感受器,主要功能是感知正羽(上文提到的片狀的羽毛,有對(duì)稱和不對(duì)稱兩種)的姿態(tài)。

纖羽
(圖片來(lái)源:allaboutbirds)

位于尾部對(duì)稱的正羽
(圖片來(lái)源:allaboutbirds)
在相關(guān)肌肉(直立肌和抑制肌,前者負(fù)責(zé)立起來(lái)羽毛,后者反之)的控制下,羽毛可以進(jìn)行“伸縮”,當(dāng)羽毛被提起來(lái)的時(shí)候,鳥(niǎo)類就會(huì)呈現(xiàn)“炸毛”狀態(tài)。一些鳥(niǎo)類頭上的羽毛就能以此表示它們受到了驚嚇。比如鳳頭鷹(一種常見(jiàn)于中國(guó)南方的國(guó)家二級(jí)保護(hù)動(dòng)物)、紅角鸮(我們俗稱“貓頭鷹”的一種)就是典型代表,在受驚時(shí)會(huì)立即豎起頭上的羽毛。

沒(méi)有立起頭冠的鳳頭鷹
(圖片來(lái)源:作者拍攝于國(guó)家動(dòng)物博物館)
不過(guò),鳥(niǎo)兒“炸毛”除了受到驚嚇,還有其它情況。比如有些是為了散熱,豎立羽毛,讓平時(shí)沒(méi)有接觸到外界空氣的皮膚不被悶著。還有的鳥(niǎo)類是為了“偽裝”,就像南美洲的栗斑翅傘鳥(niǎo)(Laniocera hypopyrra)會(huì)在幼年期晃動(dòng)自己的羽毛,假裝自己是有毒的絨蛾科(Megalopygidae)毛毛蟲(chóng)。

熱天下讓羽毛動(dòng)起來(lái)的家燕
(圖片來(lái)源:作者拍攝于北京奧森公園)

栗斑翅傘鳥(niǎo)和它偽裝出的蟲(chóng)子
(圖片來(lái)源:sci)
羽毛的適應(yīng)性轉(zhuǎn)變
對(duì)于不會(huì)飛的鳥(niǎo)而言,其羽毛的功能發(fā)生了顯著的適應(yīng)性轉(zhuǎn)變。以鸮鸚鵡和渡渡鳥(niǎo)為例,它們翅膀短小,腿部強(qiáng)健,但保留了不對(duì)稱的片狀羽毛,且初級(jí)飛羽的數(shù)目依舊是9-11根。相比之下,南方鶴鴕、大美洲鴕和小斑幾維等鳥(niǎo)類的羽毛則呈現(xiàn)出更為明顯的退化特征:不僅喪失了飛行功能,其羽片結(jié)構(gòu)也趨于簡(jiǎn)化(如鶴鴕的毛發(fā)狀羽毛與布偶貓的被毛相似),初級(jí)飛羽數(shù)目也有所改變。

大美洲鴕,羽毛看著更像毛
(圖片來(lái)源:作者拍攝于上海動(dòng)物園)
這背后的原因又是什么呢?學(xué)界對(duì)30類不飛行鳥(niǎo)類及其近緣物種的骨骼和羽毛進(jìn)行對(duì)比發(fā)現(xiàn),在飛行能力退化過(guò)程中,這些鳥(niǎo)類的腿部和翅膀演化速率更快,而羽毛形態(tài)演化速率相對(duì)慢。這是因?yàn)橥炔亢统岚蜷L(zhǎng)短對(duì)于地面生活更為重要,而羽毛生長(zhǎng)需要的能量相對(duì)骨骼與肌肉發(fā)育更低,因此演化相對(duì)滯后。羽毛的演化主要表現(xiàn)為不對(duì)稱性逐漸降低,甚至變得像小斑幾維那樣只剩下絨毛。同時(shí),翅膀上的初級(jí)飛羽數(shù)量也不再保持飛行鳥(niǎo)類典型的9-11根,而是出現(xiàn)不同程度的增減。
此外,羽毛性狀的改變還與飛行能力喪失的時(shí)間節(jié)點(diǎn)有關(guān)。在漫長(zhǎng)的演化歷程中,鳥(niǎo)類首先從有齒、長(zhǎng)尾的鳥(niǎo)翼類恐龍演化為具備飛行能力的現(xiàn)代鳥(niǎo)類,隨后部分類群又再度放棄或削弱了飛行本領(lǐng)(最早可追溯至白堊紀(jì)晚期)。其中,比起南方鶴鴕、大美洲鴕和企鵝等很早喪失飛行能力鳥(niǎo)類,較晚喪失飛行能力的鳥(niǎo)類羽毛“爆改”程度更低。
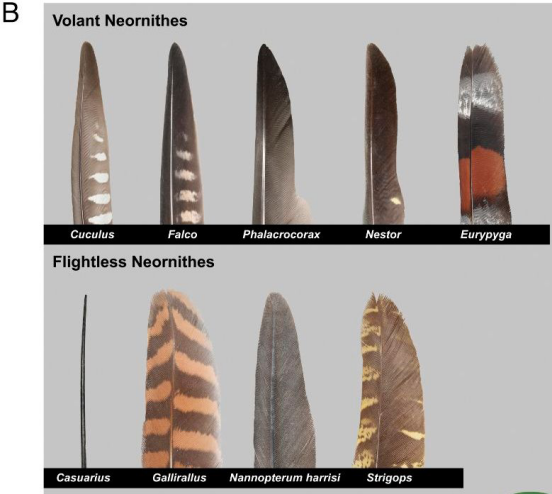
上圖為飛行鳥(niǎo)類不對(duì)稱飛羽
下圖為不飛鳥(niǎo)類的羽毛示例,其中有些留有不對(duì)稱飛羽,但是有些羽毛類似頭發(fā),如最左側(cè)的鶴鴕羽毛。
(圖片來(lái)源:參考文獻(xiàn)[12])
由此觀之,從恐龍到現(xiàn)代鳥(niǎo)類,羽毛在演化過(guò)程中展現(xiàn)出驚人的可塑性。它不僅能夠適應(yīng)飛行需求,在喪失飛行能力的類群中還表現(xiàn)出多樣化的形態(tài)與功能轉(zhuǎn)變,充分體現(xiàn)了其對(duì)不同生態(tài)環(huán)境的卓越適應(yīng)能力。
參考文獻(xiàn):
[1]Lingham-Soliar T, Murugan N. A new helical crossed-fibre structure of β-keratin in flight feathers and its biomechanical implications. PLoS One. 2013;8(6):e65849.
[2]Cincotta A, Nicola? M, Campos HBN, et al. Pterosaur melanosomes support signalling functions for early feathers. Nature. 2022;604(7907):684-688.
[3]Cooper RL, Milinkovitch MC. In vivo sonic hedgehog pathway antagonism temporarily results in ancestral proto-feather-like structures in the chicken. PLoS Biol. 2025;23(3):e3003061.
[4]O'Connor J, Clark A, Kuo PC, et al. Chicago Archaeopteryx informs on the early evolution of the avian bauplan. Nature. Published online May 14, 2025.
[5]Kiat Y, O'Connor JK. Functional constraints on the number and shape of flight feathers. Proc Natl Acad Sci U S A. 2024;121(8):e2306639121.
[6]Pan Y, Zheng W, Moyer AE, et al. Molecular evidence of keratin and melanosomes in feathers of the Early Cretaceous bird Eoconfuciusornis. Proc Natl Acad Sci U S A. 2016;113(49):E7900-E7907.
[7]Pan Y, Zheng W, Sawyer RH, et al. The molecular evolution of feathers with direct evidence from fossils. Proc Natl Acad Sci U S A. 2019;116(8):3018-3023.
[8]Padian K. 25th anniversary of the first known feathered dinosaurs. Nature. 2023;613(7943):251-252.
[9]Saitta ET, Balaji L, Mitchell JS, Makovicky PJ. Feather evolution following flight loss in crown group birds: relaxed selection and developmental constraints. Evolution. 2025;79(5):737-751.
[10]Homberger DG, de Silva KN. The role of mechanical forces on the patterning of the avian feather-bearing skin: A biomechanical analysis of the integumentary musculature in birds. J Exp Zool B Mol Dev Evol. 2003;298(1):123-139.
[11]Saitta ET, Balaji L, Mitchell JS, Makovicky PJ. Feather evolution following flight loss in crown group birds: relaxed selection and developmental constraints. Evolution. 2025;79(5):737-751.
[12]Kiat Y, O'Connor JK. Functional constraints on the number and shape of flight feathers. Proc Natl Acad Sci U S A. 2024;121(8):e2306639121.
[13]Xu X, Currie P, Pittman M, et al. Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features. Nat Commun. 2017;8:14972.
[14]Zheng X, Zhou Z, Wang X, et al. Hind wings in Basal birds and the evolution of leg feathers. Science. 2013;339(6125):1309-1312.
[15]Farzeen Shahid, Jing-Shan Zhao, Pascal Godefroit. Design of flying robots inspired by the evolution of avian flight. ARCHIVE Proceedings of the Institution of Mechanical Engineers Part C Journal of Mechanical Engineering Science .2019; 233(23): 7669 – 7686
[16]Wu P, Lai YC, Widelitz R, Chuong CM. Comprehensive molecular and cellular studies suggest avian scutate scales are secondarily derived from feathers, and more distant from reptilian scales. Sci Rep. 2018;8(1):16766.
[17] Cooper RL, Milinkovitch MC. Transient agonism of the sonic hedgehog pathway triggers a permanent transition of skin appendage fate in the chicken embryo. Sci Adv. 2023 May 19;9(20):eadg9619.

來(lái)源: 中國(guó)科普博覽
內(nèi)容資源由項(xiàng)目單位提供
