
 科普中國公眾號
科普中國公眾號
 科普中國微博
科普中國微博

 幫助
幫助
 中國科普博覽
中國科普博覽 
出品:科普中國
作者:張易、傅雄飛(中國科學(xué)院深圳先進技術(shù)研究院合成所)
監(jiān)制:中國科普博覽
動物遷徙與生物遷徙,是兩個不同的概念。我們常從紀錄片中看到的非洲大遷徙屬于動物遷徙的范疇。每年大約有超過一百萬只的角馬、數(shù)十萬只斑馬和其他草食動物從塞倫蓋蒂國家公園遷移到馬賽馬拉國家保護區(qū),尋找新的草場和水源。此外還有每年的候鳥遷徙、北美帝王蝶的遷徙等。

非洲大遷徙
(圖片來源:Veer圖庫)
而生物遷徙的概念則更加廣泛。生物遷徙是指包括動物、植物、微生物等在內(nèi)的所有生物,為適應(yīng)環(huán)境變化而發(fā)生的遷移現(xiàn)象。其中包括的植物的遷徙比如竹子等草本植物通過地下根系向外擴張形成新植株。微生物遷徙比較直觀的例子是水果表面霉斑從小變大,最后導(dǎo)致水果完全腐爛。
在生物遷徙的過程中,往往伴隨著病毒的傳播。病毒的生存方式和其他生物不同,必須依賴宿主細胞的物質(zhì)和能量才能實現(xiàn)自我復(fù)制繁殖。自然界中的病毒自身也不具備運動能力,通常需要依賴于宿主的運動實現(xiàn)傳播。
那么,問題來了,生物遷徙究竟是會促進病毒傳播,還是會抑制病毒傳播呢?這是我們今天要聊的主要話題。
傳統(tǒng)觀點:生物遷徙會加速病毒傳播
傳統(tǒng)的觀點,或者可以說是直覺上,人們認為宿主的運動會加速病毒的傳播。因為攜帶病毒的宿主運動會將病毒帶向更遠更廣的范圍,從而增加了病毒接觸新鮮宿主的概率致使病毒的傳播增強。另一方面,遷徙的宿主在遷徙的過程中會接觸到更多的病毒,從而導(dǎo)致感染病毒的幾率變大。
例子一“科學(xué)家Figuerola曾在2000年分析了雁形類比如雁、鴨、天鵝等遷移中寄生蟲感染的多樣性和流行度,數(shù)據(jù)表明每個宿主身上的寄生蟲種類或者屬數(shù)與遷徙距離成正相關(guān),在經(jīng)歷較長距離遷徙的物種中,更高比例的個體被寄生蟲感染[1]。

大雁遷徙
(圖片來源:Veer圖庫)
例子二:1999年至2000年期間,西尼羅河病毒最初沿著北美鳥類遷徙的主要走廊蔓延,并迅速從其在大西洋海岸的紐約市起源點向外擴散[2]。盡管雀形目鳥類候鳥是西尼羅河病毒的適宜宿主并且能夠作為有效的傳播媒介,但對病毒擴張是由候鳥的活動造成的這個定論科學(xué)界依然是模棱兩可的。另一項研究表明,在2007年期間,剛果民主共和國每年遷徙果幅的涌入時間與當?shù)卮迩f人類埃博拉疫情爆發(fā)的開始時間相重合[3]。
遷徙淘汰假說:生物遷徙會抑制病毒傳播
“遷徙淘汰”假說沒有明確的提出者,它的形成和發(fā)展與多個科學(xué)家的研究密切相關(guān)。
最初,周期性遷徙被認為是一種減少寄生蟲出現(xiàn)的宿主策略,但提出的最主要機制是逃避在宿主周圍環(huán)境中累積的寄生蟲。然而,種群遷徙的過程也可能將患病動物淘汰,從而保持了遷徙群體的健康性。
20多年來,美國生態(tài)學(xué)家Altizer一直以帝王蝶為模型研究季節(jié)性遷徙對寄生蟲流行率的影響,發(fā)現(xiàn)遷徙群體中確實發(fā)生了遷徙淘汰現(xiàn)象。他們對三種不同遷徙類型的帝王蝶持續(xù)追蹤,北美東部的帝王蝶A種群每年秋天從加拿大以北的地區(qū)遷徙多達2500公里到達墨西哥中部的過冬棲息地。北美西部的帝王蝶B種群遷徙距離較短,在加利福尼亞海岸附近過冬。而佛羅里達南部的帝王蝶C種群是非遷徙種群世代在此生活。

帝王蝶(大閃蝶)
(圖片來源:Veer圖庫)
這三種不同遷徙類型的帝王蝶,研究人員通過野外研究和實驗研究發(fā)現(xiàn)[4],感染了寄生蟲的帝王蝶飛行距離較短,飛行速度也較慢,在秋季遷徙期間,隨著帝王蝶向南移動,寄生蟲的流行率降低,這與“遷徙淘汰”理論一致。而“遷徙淘汰”又導(dǎo)致了不同遷徙行為的帝王蝶中寄生蟲流行率的顯著差異:長距離遷徙的A種群的寄生蟲流行率低于短距離遷徙的B種群,而非遷徙C種群的寄生蟲流行率最高。并且研究還表明,長距離遷徙種群A的寄生蟲分離株的毒性低于短距離遷徙種群B和非遷徙種群C的分離株,這說明較長的遷徙距離會淘汰攜帶高毒性寄生蟲基因型的帝王蝶。
兩種觀點爭執(zhí)不下,到底真相如何呢?
兩種觀念爭議的焦點主要是生物的遷徙運動到底是如何影響病毒傳播的,或者說是什么模式的遷徙運動會促進/抑制病毒的傳播?并且在病毒“遷徙淘汰”的機制猜想上,以往的研究處于無法統(tǒng)一的狀態(tài),比如有些學(xué)者認為是病毒感染導(dǎo)致遷徙能力的下降從而導(dǎo)致了“遷徙淘汰”,例如比尤伊克天鵝(cygnus columbianus bewickii)的研究表明,禽流感病毒的感染降低了受感染鳥類的飛行距離,但對白額雁禽流感病毒的研究又并未發(fā)現(xiàn)受感染鳥類和未受感染鳥類的遷徙距離有任何差異。
所以宿主的運動是如何影響病毒傳播的呢?在什么模式下宿主運動抑制病毒傳播?而抑制狀態(tài)下的“遷徙淘汰”機制到底是什么?這是我們團隊(中國科學(xué)院深圳先進研究院研究員傅雄飛團隊)感興趣并且期望探明的問題。
追蹤動物遷徙耗時耗力,我們設(shè)計了新思路!
以往的生態(tài)學(xué)家追蹤動物遷徙及其中的病毒流行率主要依賴于田野調(diào)查,比如在動物遷徙的停留點捕獲一定數(shù)量的動物收集其血液、糞便、唾液等樣本以監(jiān)測病原蟲或病毒的種類及數(shù)量,然后將其放回自然界種,在它們遷徙的未來線路上再次捕獲并記錄,從而觀察動物生理狀況及病原蟲豐度的變化。
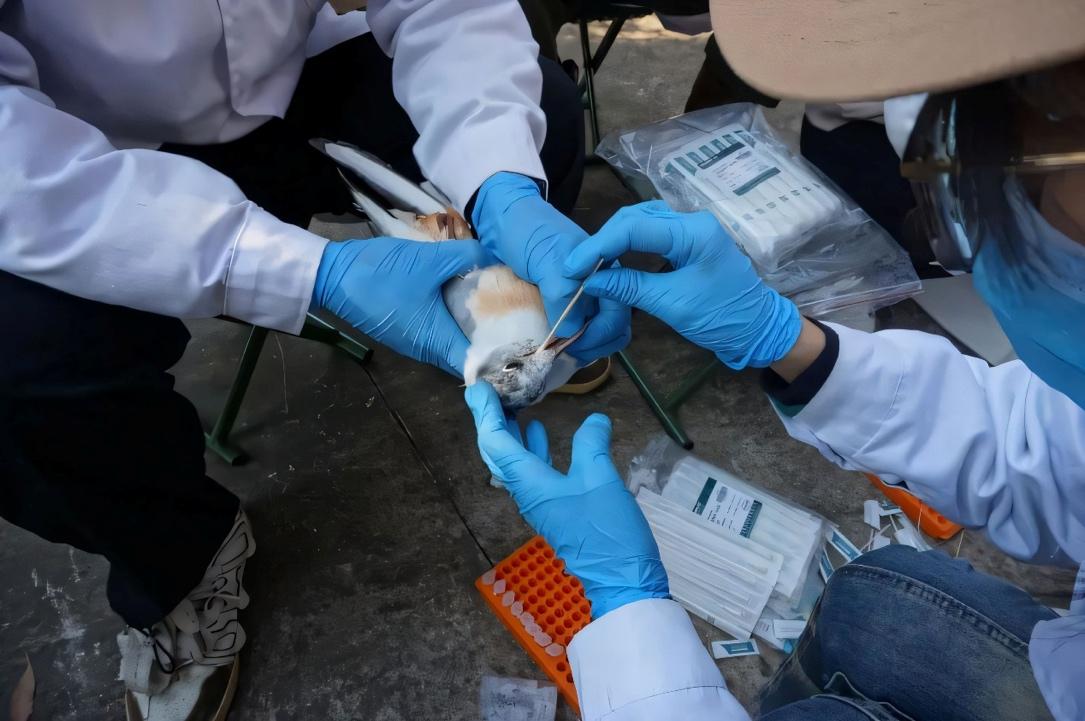
開展紅嘴鷗環(huán)志與疫病監(jiān)測
(圖片來源:昆明市林草局)
整個過程需要耗費大量的人力物力,樣本數(shù)量有限,往往最終統(tǒng)計的數(shù)量達不到期望值。并且,自然界環(huán)境更復(fù)雜多變,研究人員無法控制研究過程中的變量,從而導(dǎo)致在分析結(jié)果的時候無法歸結(jié)于某個單一關(guān)鍵因素。此外,以往針對病毒傳播時空動力學(xué)的理論也大多屬于總結(jié)的經(jīng)驗性理論,缺乏實驗體系的驗證。
于是,我們團隊引入了合成生物學(xué)的手段,想到了一個新思路:在實驗室模擬一場生物大遷徙,讓我們不出門也能開展追蹤研究!
細菌是自然界分布最廣個體數(shù)量最多的有機體。而噬菌體是一類細菌依賴性的病毒,具有高度的宿主特異性,僅寄居在特異的宿主菌體內(nèi)。噬菌體侵入細菌細胞后,利用宿主細胞內(nèi)物質(zhì)和能量進行自身的復(fù)制繁殖,產(chǎn)生子代噬菌體,子代噬菌體釋放出宿主體外后又會繼續(xù)感染新的細菌。細菌-噬菌體的互相作用過程是一個天然的病毒傳播問題。并且,現(xiàn)在合成生物學(xué)技術(shù)的成熟又可以讓科研人員隨意調(diào)控噬菌體和宿主細菌的各項功能元件,因此,噬菌體和宿主細菌的相互作用為研究流行病學(xué)中病毒的傳播問題提供了一種很好的實驗定量研究體系。
因而,我們選擇了大腸桿菌這種實驗室里司空見慣并且又被研究的很透徹很容易被基因編輯調(diào)控的細菌作為宿主細菌,選擇M13這個可特異性感染大腸桿菌的溫和型噬菌體作為病毒,構(gòu)建了一個實驗室下的宿主病毒空間互相作用實驗體系,來研究宿主運動如何影響病毒傳播這個生態(tài)學(xué)問題。
具體我們是這樣操作的:大家可以想象一下橙子表面的霉斑,從一個小霉斑點慢慢向外擴張生長成一個大霉斑。我們做的實驗很類似,我們將一小滴大腸桿菌液體接種到內(nèi)含凝膠狀營養(yǎng)物質(zhì)的培養(yǎng)皿上之后,細菌會迅速地繁殖然后向外進行空間擴張。具體的過程就是擴張的最前沿細菌群體向外運動占領(lǐng)新的地盤,然后一部分細菌駐扎下來繁殖更多后代從而徹底占領(lǐng)此地,與此同時,另一部分細菌繼續(xù)向外運動征戰(zhàn)新的地盤。因而這個擴張前沿的細菌群體是一個天然的群體遷徙系統(tǒng)。
如果我們在菌液外一定距離的地方放置一滴噬菌體病毒液體。當空間擴張的細菌群體碰上了噬菌體,就會迅速的被噬菌體感染,隨后被感染的細菌群體一方面跟隨大部隊向前繼續(xù)擴張,另一方面,他們開始產(chǎn)生新的噬菌體然后感染他們相鄰的健康細菌。而固定位置處擴張前沿遷徙群體中噬菌體感染者變化的速率就可以表征遷徙群體內(nèi)病毒的傳播能力。基于這個思路,我們利用合成生物學(xué)技術(shù),通過調(diào)控宿主細菌的運動性以及病毒的繁殖能力,就可以研究宿主的運動性是如何影響病毒傳播的。
終于,我們有了新的發(fā)現(xiàn)
最終,我們發(fā)現(xiàn):像燈光可以吸引飛蛾一樣,當化學(xué)物質(zhì)對細菌具有吸引性時(即趨化性;該狀況下的細菌空間擴張被稱之為定向型空間擴張),細菌的空間擴張速度越快,擴張前沿中噬菌體的傳播能力越弱。反之,化學(xué)物質(zhì)對改造后的細菌沒有吸引性時(該狀況下的細菌空間擴張被稱之為無方向型空間擴張),改造的細菌空間擴張速度越快,擴張前沿中噬菌體的傳播能力反而越強。
我們構(gòu)建了細菌-噬菌體空間互相作用的數(shù)學(xué)模型,成功重現(xiàn)了細菌運動性對病毒傳播可促進可抑制這個截然相反的現(xiàn)象。究其原因,我們發(fā)現(xiàn)是宿主對化學(xué)物質(zhì)的趨化性運動是抑制病毒傳播的關(guān)鍵因素。我們利用合成生物學(xué)技術(shù)通過調(diào)控對化學(xué)物質(zhì)的敏感度從而改變細菌的趨化運動能力,成功驗證了細菌的趨化運動抑制病毒的傳播。
此外,我們還發(fā)現(xiàn),當噬菌體的繁殖能力比較弱的時候,增大細菌的定向遷移速度,病毒甚至能從整個擴張前沿中消失,也即是發(fā)生了“遷徙淘汰”現(xiàn)象。我們對該現(xiàn)象追根究底探索發(fā)現(xiàn),引發(fā)“遷徙淘汰”的最根本原因是因為病毒不會運動,被病毒感染的細菌產(chǎn)生的子代病毒留在原地,它只能感染原地位置臨近處的新宿主,這就導(dǎo)致最前沿的遷移群體中健康的宿主位于前方,而被感染的病毒位于后方,健康者和感染者在空間上存在前后有序的空間分布差異。然而,擴張前沿的細菌群體在向前運動的時候會遺留一部分群體駐扎下來繁衍生息徹底占領(lǐng)該位置,因而位于擴張前沿中偏后方的被感染宿主會更容易被淘汰出去,并且遷徙的速度越快,這個淘汰率越大,當速度大到一定程度時,被感染宿主就被完全的清除出擴張前沿了,也即是發(fā)生了“遷徙淘汰”。
當然,現(xiàn)實自然界中的環(huán)境更復(fù)雜,前文提到的被感染宿主生理機能變差等因素會進一步加劇遷徙淘汰現(xiàn)象,但我們的研究提供了對遷徙淘汰現(xiàn)象的另一種新解讀,研究成果最終發(fā)表在了美國《國家科學(xué)院院刊》上。
這一研究成果對于流行病防控等具有重要意義
盡管現(xiàn)實中存在更為復(fù)雜和多變的人為及環(huán)境因素,這項研究為解釋遷徙型的北美帝王蝶中攜帶病原蟲的比例遠低于定居型的帝王蝶,以及生態(tài)學(xué)中其他類似現(xiàn)象——即群體遷徙可能導(dǎo)致種群中病毒豐度下降——提供了全新的視角。
只有理解病毒在這些遷徙群體中傳播的動力學(xué)特征,人類才能預(yù)測野生動植物和人類未來的傳染病風(fēng)險。
“細菌-噬菌體共遷移中自發(fā)形成的感染者位于后方,而健康者位于前方的這種空間有序分布,致使感染者更容易被淘汰”的這一機制提示:在流行病學(xué)背景下制定防控策略時,可以綜合考慮生態(tài)與行為因素的相互作用,并通過調(diào)控生物遷移行為或病毒感染特性等關(guān)鍵因素,應(yīng)對傳染病的傳播。
例如,在植物的自然擴張中存在病蟲災(zāi)害時,因其行為屬于非方向性的空間擴張,該種模式下宿主運動是促進病毒傳播的,因而要人為干預(yù)及時清理病蟲植株以防病蟲災(zāi)害。而對于類似帝王蝶等存在明確目標的物種空間擴張中,宿主運動會抑制病毒傳播,在防控帝王蝶群體中病毒的傳播策略時則可適度降低干預(yù)。
除了指導(dǎo)非致命性傳染病防控,該研究也為其他如致命病毒在宿主中的傳播問題提供了新的策略和實驗手段,例如可通過在實驗室體系下構(gòu)建“宿主-致死性病毒”互相作用實驗系統(tǒng)定量系統(tǒng)地研究致命性病毒傳播問題。
并且,該研究通過在分子層面定量調(diào)控細菌遷移速度和噬菌體侵染能力,結(jié)合數(shù)理模型,揭示了在定向空間擴張中宿主與病毒共遷移系統(tǒng)的時空有序結(jié)構(gòu),從而解釋了群體層面生物的“遷徙淘汰”機制,實踐了定量合成生物學(xué)“造物致知”的理念。
定量合成生物學(xué)作為一項前沿交叉學(xué)科,該研究為物理學(xué)、定量生物學(xué)、計算生物學(xué)、生態(tài)學(xué)和傳染病學(xué)等多學(xué)科的交叉融合提供了示范。
下一步,我們團隊計劃在實驗室體系下構(gòu)建“宿主-致死性病毒”互相作用的合成生態(tài)實驗系統(tǒng)定量系統(tǒng)地研究致命性病毒傳播問題,從而對現(xiàn)實世界中傳染病傳播的問題進行更深入全面的研究。
合成生物學(xué)是一個快速發(fā)展的領(lǐng)域,我們相信,未來,合成生物學(xué)在疾病治療、生物制造、環(huán)境保護、合成生命、合成生態(tài)系統(tǒng)等領(lǐng)域的突破,不僅有助于推動科學(xué)技術(shù)的發(fā)展,還將對社會、經(jīng)濟和環(huán)境產(chǎn)生深遠的影響。
參考文獻:
1. J. Figuerola, A. J. Green, Haematozoan Parasites and Migratory Behaviour in Waterfowl. Evolutionary Ecology 14, 143–153 (2000).
2. J. H. Rappole, S. R. Derrickson, Z. Hubálek, Migratory Birds and Spread of West Nile Virus in the Western Hemisphere. Emerg. Infect. Dis. 6, 397–400 (2000).
3. E. M. Leroy, et al., Human Ebola Outbreak Resulting from Direct Exposure to Fruit Bats in Luebo, Democratic Republic of Congo, 2007. Vector-Borne and Zoonotic Diseases 9, 723–728 (2009).
4. S. Altizer, R. Bartel, B. A. Han, Animal Migration and Infectious Disease Risk. Science 331, 296–302 (2011).

來源: 中國科普博覽
內(nèi)容資源由項目單位提供
