
 科普中國公眾號
科普中國公眾號
 科普中國微博
科普中國微博

 幫助
幫助
 科普中國北大行之“人與自然和諧共生”
科普中國北大行之“人與自然和諧共生” 
天生的方向感——鳥類的指南針是怎么工作的?
從“木落雁南度,北風江上寒”到“萬里人南去,三春雁北飛”,鴻雁南渡北歸的身影穿梭于千年前的詩詞文脈里,是古人對候鳥長距離遷徙的描述與驚嘆;無論是漢代蘇武牧羊中鴻雁傳書的典故,還是唐代張九齡宰相飛鴿傳信的軼事,都反映出鳥類在另一種歸巢任務中同樣精準的定向能力。
其實,強大的定向導航能力不只見于大雁和鴿子,而是許多鳥類都具備的特質:每年,數十億只小型鳴禽離開北極和溫帶繁殖區,跨越數千公里的距離在熱帶和亞熱帶地區越冬。鳥類當中不乏卓越的遠航者:斑尾塍鷸 (Limosa lapponica)從阿拉斯加飛往新西蘭的旅程持續 7-9 天,從不停歇;北極燕鷗 (Sterna paradisaea) 在北極附近繁殖,在南極附近越冬,其航程跨越整個星球;還有海鷗,它們每年飛行超過十萬公里,只為返回浩瀚海洋中的小島繁衍生息[1]……從古至今,無論個例還是廣例,無不展現著鳥類導航能力的強大。

圖一 斑尾塍鷸 (Limosa lapponica)

圖二 北極燕鷗 (Sterna paradisaea)
圖源網絡
面對如此強大的定位能力,人類固有的優越感蕩然無存。同樣是脊椎動物,共享同樣的視聽覺感受機制,甚至擁有不及人類拳頭大小的腦,鳥類天生的方向感究竟從何而來?除了更高分辨率的常規感知之外,唯一可能的理由,就是它們具備人類不配備的特殊感覺系統。
那么,到底是什么特殊的地球物理量蘊含著精度極高的位置和方向信息,服務于鳥類的遠程飛行?經過數個世紀的研究,科學家最終把目光鎖定在一類特殊的物理量——地磁場——之上。
地磁場是地球表面及近地空間中存在的天然磁場。如圖,磁場的強度由磁力線的疏密表征,在兩極地區最大,在赤道地區最小。磁場形狀類似條形磁鐵的偶極子場,磁場的方向可以由磁力線方向表示,從南半球出發,跨越地球表面,在北半球重新進入地球內部。磁力線和地球之間形成的傾角隨緯度而變化——在磁赤道處,磁力線平行于地球表面,傾角為 0°;從磁赤道向兩側,傾角逐漸陡峭,磁極處磁力線直接向下指向地球,傾角為 90°[2]。場強和傾角均可以編碼位置信息——由于不同區域的磁場強度存在差異,因此前者可起到“地圖”的作用;后者則攜帶方向信息作為“指南針”。

圖三 地球磁場[2]
對于鳥類利用磁場導航的可能性早在19世紀就被提出和討論。在Alexander von Middendorf關于俄羅斯鳥類的論文中指出:“......候鳥驚人的一心一意——不顧風和天氣,不顧黑夜和霧——可能是因為家禽總是知道磁極的方向,因此也知道如何準確地保持它的方向”[3]。
動物感知地磁場能力的存在這一觀點具有革命性意義,但直到20世紀中期才具備充分的技術條件加以檢驗。20世紀50年代,通過外加亥姆霍茲線圈干擾定向遷徙的行為學實驗,科學家首先證明了一種夜間遷徙的鳴禽,歐洲知更鳥(Erithacus rubecula),具備磁場導航的能力[4]。
在隨后的幾十年里,絕大部分鳥類的磁場導航能力都得到了驗證;有意思的一點是,鳥類磁定向系統需要依賴光照發揮作用:在可透弱光的木箱中長距離運輸鴿子,隨后記錄其歸巢方向選擇,發現當在木箱周圍施加隨機干擾磁場時,鴿子的確完全失去了定位能力;而將箱子完全隔絕光照,也能起到與干擾磁場一致的效果,證明鴿子的磁定向能力依賴于光照的存在,即使是很弱的光強[5]。
磁感受依賴光照的特殊性質,意味著鳥類似乎能“看”到磁場;換言之,視覺系統與磁場感知密切相關。然而,磁場信息是如何經歷一系列物理化學變化轉化為神經信號的?近一個世紀以來,經過無數的假設與檢驗,鳥類磁感覺的神秘面紗正在被一點點揭開。
目前領域內主流的觀點認為,磁場的強度和方向分別被兩個正交的感覺系統編碼:前者會通過激活鳥類頭部含有磁鐵礦晶體的感受器細胞,使鳥類感知磁場強度進而得出位置信息,與光照關聯不大;后者則通過視覺系統起作用,以一種奇妙而罕見的自由基對機制影響視網膜上的化學反應方向,最終使鳥類得以感知磁場方向。在此我們將主要探討后一種機制——鳥類是如何“看”到磁場方向的?
自由基對參與鳥類磁場感知的理論可以追溯到 1978 年,Schulten 等提出磁敏感光化學轉變的假說,自由基對作為反應中間體,化學反應產物的產率取決于地磁場相對于反應物分子的方向[6]。
乍一看,這種理論幾乎難以置信:在生理條件下,地球磁場(30-65 μT)與單個分子的相互作用比其熱能k B T小一百萬倍以上,很難對熱化學反應動力學或熱力學產生影響。然而,不妨設想這樣一個場景:有一個沉重的石塊,蒼蠅是否能夠通過撞擊來使其傾倒?答案顯然是否定的。但是,如果先給予一定的能量,使石頭僅靠一條棱保持平衡。此時即使蒼蠅撞擊傳遞的能量很小,也足以導致塊落到右側而不是左側。
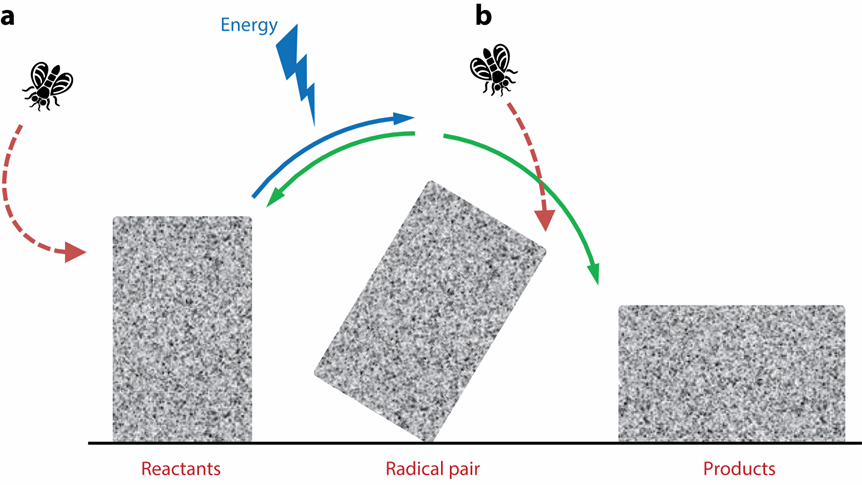
圖四 自由基對機制的類比模型[7]
在化學反應熱力學系統中,平衡態均處于能量谷,不同之處在于前者是穩定平衡,位于能量最低的谷;后者則屬于不穩定平衡,能量谷的相對值高于反應物和產物,微小的擾動即可使系統偏離本來狀態,跌至能量更低的另一個谷。自由基對機制就是利用非平衡態系統的不穩定性發生的反應:非平衡態是自由基對,達到該狀態所需的能量來自光子。
那么,反應的最佳載體是什么呢?常規的哺乳動物光感受器視蛋白將光能轉化為一種視覺信號轉導分子(視黃醛)的構象變化,不涉及自由基對反應;然而對鳥類視覺系統的研究表明,候鳥的眼睛中還存在隱花色素,且其種類多達四種[8],具有發生自由基對反應的潛力。
接下來,我們就簡要介紹一下自由基對反應機理及其實現磁場感知的機制,并探討隱花色素是如何介導自由基反應發生的。
一、自由基對反應機理
我們知道物質由原子組成,原子又由原子核圍繞電子形成。穩定狀態下,電子總是以兩兩成對的形式存在,但化學反應中有可能產生包含未成對電子的物質,也就是自由基。一個自由基對由兩個自由基組成,這兩個自由基通常是通過單分子均裂或者雙分子間電子轉移產生的。例如甲烷(CH4)總共有10個電子,其中 6 個由碳貢獻,4個來自氫。兩個內層電子圍繞著碳原子核,八個外層電子參與形成碳氫鍵,每個鍵兩個電子。如果其中一個鍵均裂,得到兩個電中性基團:甲基自由基 CH3 ? 和氫原子H?。
由于電子具有自旋角動量的特性,類似一個繞其軸旋轉的小球形物體,因此當電子帶電旋轉,會產生一個磁場,就像線圈中的電流,因此電子可視作一個微觀磁鐵,具有一個固定磁矩。CH3 ? 有 9 個電子,其中 8 個成對存在,因此它們的磁矩正好抵消。因此,自由基的磁性源于未成對電子。
電子自旋角動量的特殊之處在于其具有量子特性,大小由自旋量子數 S 決定,單位為±1/2。量子力學規定,一個自由基對如CH3 ? 和H? 的自旋量子數只能為 0 或 1,可以理解為兩個不成對電子的自旋可以平行(↑↑, S=1)或反平行(↑↓,S=0)。由于理論認為自旋量子數為S的物質有2S+1種態,故S = 1為三重態,S=0為單重態;因此,自由基對的兩種形式稱為三元組(S=1) 和單元組(S=0)。
自由基對反應中,電子必須自旋守恒。由于自由基反應的產物為單元組的分子,因此最直接的中間體自由基必須也為單元組,而三元組自由基則由單元組自由基轉化而來。
此外,自由基對還具有兩個顯著的特征:其一,存在廣泛的超精細相互作用,可以簡單理解為自由基中離域的不成對電子同時與多個原子核相互作用,且這種作用是不對稱的(隨角度變化),構成了磁場方向感知的基礎;其二,由于量子力學原因,單重態和三重態之間存在高頻相互轉化,也即單重態分子的占比處于高頻振蕩中(圖五a)。這一震蕩是由內部原子核自旋引起的,也因此,這一震蕩很容易受到外部磁場波形的調制:不同頻率的波疊加,形成低頻包絡波形和高頻振蕩波形。而由于超精細相互作用的不對稱性,不同方向的磁場會產生不同的疊加波形(圖五b、c)。
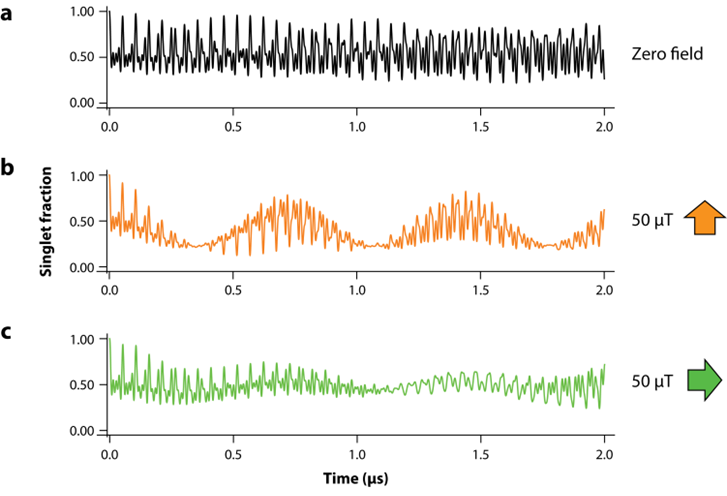
圖五 單重態自由基分數隨時間變化情況[7]
基于上述的自由基對反應特征,一個精致的模型應運而生(圖六a):光激發下,一個單重態自由基對瞬時形成,成為兩個相互競爭的反應的原料:其可以直接或間接(轉化為三重態)生成產物,也可以通過逆反應回到反應物。
如果與外場的相互作用增加了平均三重態分數(單重態分數相應減少),那么更少的自由基對將恢復回 AB,更多的自由基對生成 C 做出貢獻,反之亦然。因此C 的最終產率取決于外部磁場的存在和方向。這就是磁場效應的起源。盡管自由基對振蕩對于磁場效應的存在至關重要,但一旦所有自由基對都消失,C 的最終產率將為鳥類提供有關磁場方向的信息。
這個模型可以做如下類比(圖六b):兩個標有 S 和 T 的浴缸由一根帶閥門的管子連接,其中的水量分別對應于單重態和三重態自由基對的分數。T 在底部有一個出口,該出口排空到標記為 C 的桶中,而 S 有兩個出口,其中一個通向 C,另一個通向AB。初始情況, S 裝滿了水,T、AB、C 為空,瓣膜部分打開。水從 S 流到 T,同時落入桶中,直到浴缸排空。最終進入兩個桶中的水量(AB 和 C 的最終產量)取決于管子的直徑(速率常數k S 和 k C 的值)以及將閥門轉動的程度(外加磁場對轉化情況的調制)。如果閥門完全打開,使水從 S 快速流向 T,則有兩條有效的路徑到達 C;如果閥門部分關閉,則到達 T 的水就會減少,到達 C 的唯一有效路線是直接從 S 到達,最后C 中的水比閥門完全打開時少。
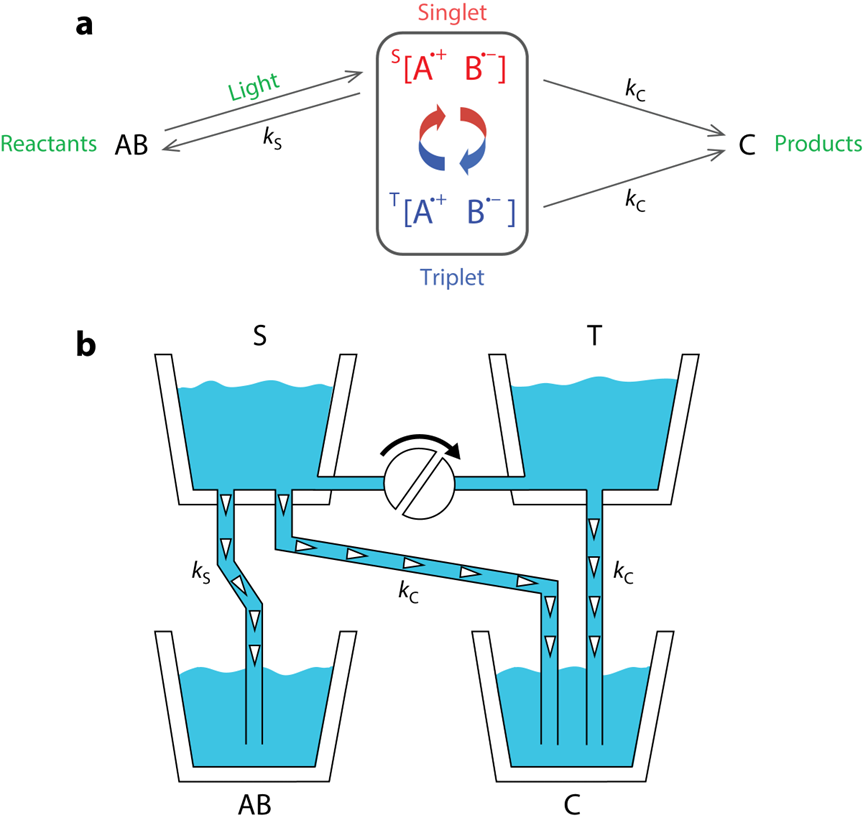
圖六 自由基對反應機理示意圖[7]
二、隱花色素參與磁場方向感知
隱花色素,顧名思義,是最早在植物中發現的一種感光色素,參與植物生長發育的光依賴性調節。其有一個結合域非共價結合FAD(氧化還原活性黃素腺嘌呤二核苷酸輔因子,其還原態FADH參與呼吸作用電子傳遞鏈的重要一環,其氧化態FAD則可以吸收藍光)。FAD 的光還原是通過色氨酸(Trp,W) 殘基三聯體的三個連續電子轉移介導的,生成黃酮半醌自由基 FAD??和TrpH+?(圖七a)。
進一步的生化實驗證實了隱花色素參與自由基對反應機制,自由基對中間體命名為RP1(radical pair1),而產物C對應次級自由基,命名為RP2(圖七b)。
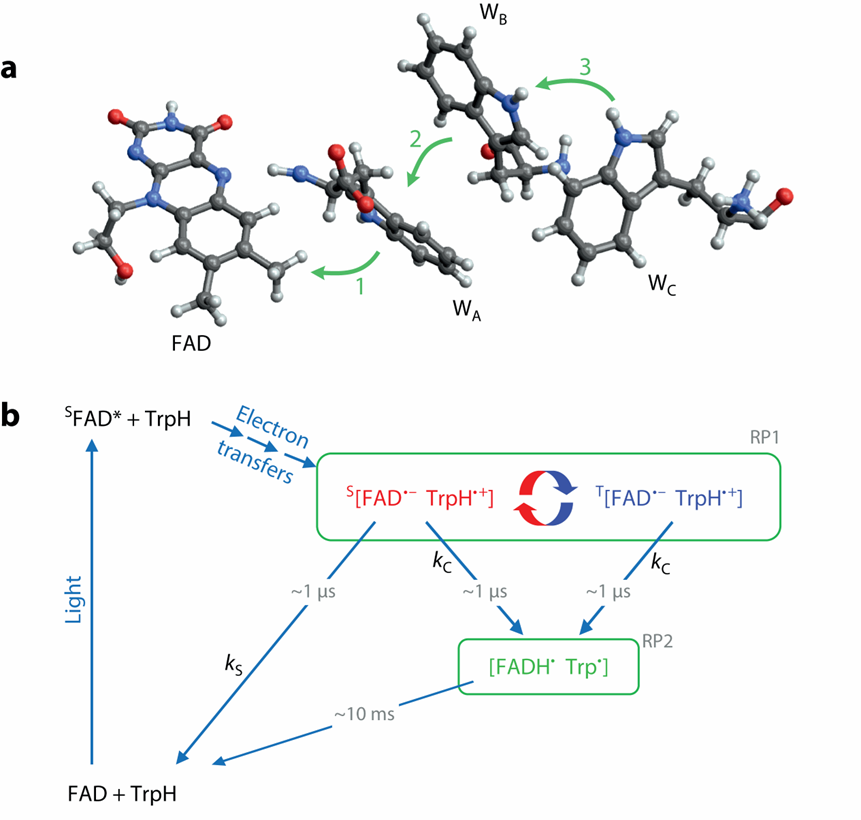
圖七 隱花色素參與自由基對反應的化學機理[7]
至此,化學家證明了自由基對反應介導磁場方向感應的可能性,并證明隱花色素是一種很有希望的候選分子。然而事實是否如此?這個問題需要由生物學家回答。
有研究表明,在夜間活躍的候鳥中,磁取向行為期間的神經活動與視網膜神經節細胞中的隱花色素表達共定位,而在非候鳥或白天沒有這種共定位[9]。遺憾的是,目前尚缺乏更為直接的證據,只能證明隱花色素參與磁場感知,但不能排除它在磁場感知的上游或下游起間接作用的可能性。此外,行為學實驗發現磁感受的機制只存在于右眼:被測試的遷徙鳥類被遮住左眼時,定向能力并未出現顯著下降,而被遮住右眼時未能表現出定向行為(圖十二)[10]。未來需要更多操縱性的實驗建立起從分子、細胞到環路和行為的系統研究。
看似微不足道的磁場能量,通過感光色素的自由基反應巧妙調控了物質的產率,以千分之一秒的精度在鳥類的視網膜上書寫精確而具體的磁場信息——行文至此,不知你是否和我一樣,沉思并且贊嘆,感慨自然選擇的神奇和時間的強大。
仰望天穹,碧藍的寰宇襯托出飛鳥纖細的身姿,它們眼中是否有一條條磁力線綿延而過?或許化學家和生物學家永遠不能回答這個問題;但至少在某一點上,我們和千年前的古人達成了共識——遠航的終點,是家的方向。
參考文獻:
[1]Mouritsen, Henrik. “Long-distance navigation and magnetoreception in migratory animals.” Nature vol. 558,7708 (2018): 50-59. doi:10.1038/s41586-018-0176-1
[2]Johnsen, S?nke, and Kenneth J Lohmann. “The physics and neurobiology of magnetoreception.” Nature reviews. Neuroscience vol. 6,9 (2005): 703-12. doi:10.1038/nrn1745
[3]von Middendorff A. Die Isepiptesen Ru?lands. Mém Acad Sc. S Petersbourg VI Ser Tome. 1859;8:1–143.
[4]Wiltschko, W. (1968), über den Einflu? statischer Magnetfelder auf die Zugorientierung der Rotkehlchen (Erithacus rubecula). Zeitschrift für Tierpsychologie, 25: 537-558. https://doi.org/10.1111/j.1439-0310.1968.tb00028.x
[5]Wiltschko, W., Wiltschko, R. Disorientation of inexperienced young pigeons after transportation in total darkness. Nature 291, 433–434 (1981). https://doi.org/10.1038/291433a0
[6]Schulten, K., Swenberg, C. & Weller, A. (1978). A Biomagnetic Sensory Mechanism Based on Magnetic Field Modulated Coherent Electron Spin Motion. Zeitschrift für Physikalische Chemie, 111(1), 1-5. https://doi.org/10.1524/zpch.1978.111.1.001
[7]Hore, P J, and Henrik Mouritsen. “The Radical-Pair Mechanism of Magnetoreception.” Annual review of biophysics vol. 45 (2016): 299-344. doi:10.1146/annurev-biophys-032116-094545
[8]Nie?ner C, Gross JC, Denzau S, Peichl L, Fleissner G. 149. et al. 2016. Seasonally changing cryptochrome 1b expression in the retinal ganglion cells of a migrating passerine bird. PLOS ONE 11:e0150377
[9]H. Mouritsen, U. Janssen-Bienhold, M. Liedvogel, G. Feenders, J. Stalleicken, P. Dirks, R. Weiler, Cryptochromes and neuronal-activity markers colocalize in the retina of migratory birds during magnetic orientation, Proc. Natl. Acad. Sci. U.S.A.101 (39) 14294-14299,https://doi.org/10.1073/pnas.0405968101 (2004).
[10]Wiltschko, Wolfgang et al. “Lateralization of magnetic compass orientation in a migratory bird.” Nature vol. 419,6906 (2002): 467-70. doi:10.1038/nature00958
來源: 科普中國北大行之“人與自然和諧共生”
內容資源由項目單位提供
