
 科普中國公眾號
科普中國公眾號
 科普中國微博
科普中國微博

 幫助
幫助

最近,就讀于某植物研究所的準碩士三年級研究生小美有點苦惱。她茶飯不思,甚至連實驗室的離心機都沒心情搶。原來,前一天晚上,隔壁實驗室的小帥向她表白了,還約她三天后去吃燒烤(即約會)。
小美對小帥不太熟,實驗室畢竟隔墻如隔山。她只能通過自己嚴謹的思維與全方位的綜合考慮,做出“接受”OR“拒絕”的決定:
接受→她可能不得不放棄寶貴的實驗時間。眾所周知,小美馬上就研三了,是人生又又又又一個關鍵時期,稍有延誤,實驗無法按時完成、文章沒法接收、延期畢業、不能按時考博、博士不能按時畢業,最終找不到工作、失去年齡優勢、申請不到基金、退休前評不到正高,完啦!科研道路全毀了!
拒絕→她可能沒法在30歲前找到對象、35歲前生不了孩子、孩子沒法在她退休前獨立,完啦!人生規劃全毀了!況且,不能落入經驗主義的圈套中,要用理性和發展的眼光看問題,維持現狀并不一定是最好的。萬一小帥實驗技能卓越,拒絕他豈非錯失一臺多功能全自動“點樣機”?
小美躊躇不已,然而三天內就要做出回復。于是她決定采取一種古老但科學有效、同時符合她植物學研究生身份的方法來做決定:數花瓣——單數接受、雙數拒絕。
小美快速地制定了實施方案,唯一的問題是:要用什么植物的花來數呢?
首先,要明白“數花瓣”這件事的本質——“花瓣數量的奇偶性”。
植物的花瓣數量是否有規律呢?在回答這個問題之前,先明確一下“花瓣”是什么?
裸子植物沒有真正的花,花是被子植物的生殖器官。一朵比較典型的花可以分成幾個部分,除了花梗和花托,從外到內直觀的有:花萼、花冠(單個花瓣稱為花瓣,所有花瓣的總和稱為花冠)、雄蕊群和雌蕊群。這些花部分之間有時候涇渭分明,有時候又模糊不清。

有時花萼和花瓣難以區分,索性也就不區分,直接合稱為花被片;有時候花瓣會長成一桶像連衣裙的東西,基部連在一起,稱為合瓣花,頂上分開的部分,稱為裂片;有時候我們以為的花瓣并不是花瓣,而是其他的東西,比如說三角梅的“花瓣”其實是苞片;有時候花瓣數量很難說清,比如說向日葵是一堆小花長在一起拼成的一個頭狀花序,如果統計小花的花瓣數量,那么向日葵周圍每朵小花只有一片花瓣(舌狀花)、內部的每朵小花有4-5個裂片(管狀花),我們是數哪種小花的花瓣數量?抑或數整個花序的花瓣數量?




為了避免小帥過后申請復查,數花瓣就要數地地道道的花瓣,哪怕小帥找公證處來也只能承認是花瓣:1)合瓣花,數裂片數量;2)無法明確區分花瓣花萼、但是看上去像花瓣的(柔軟、非綠色、光滑、不像葉子),全當花瓣數;3)如果花瓣和花萼有區分,但是中間有過渡狀態,這種花不用于計數;4)三角梅和向日葵這樣“曖昧”的花不用于計數;5)像水稻那樣,花瓣退化或缺失而難以辨認的花不用于計數(沒人想數水稻花吧)。
技術路線A:自然界有規律的花瓣數量
花是進化來的。起初,植物并沒有花,也沒有花瓣,后來葉子發生了變異,形成了花瓣,花瓣數量也是植物進化的結果。因此花萼、花瓣、雌雄蕊的著生方式同葉的著生方式有異曲同工之處——被子植物葉片有螺旋和輪生等著生方式,花器官也有螺旋和輪生。要注意的是,螺旋花不一定所有部分都是螺旋的,比如說木蘭科,它們的雄蕊螺旋著生,但花被片輪生;睡蓮目的花瓣螺旋著生,但萼片常常只有1輪(3-5枚左右)。目前還說不清到底是螺旋花更原始還是輪生花更原始。

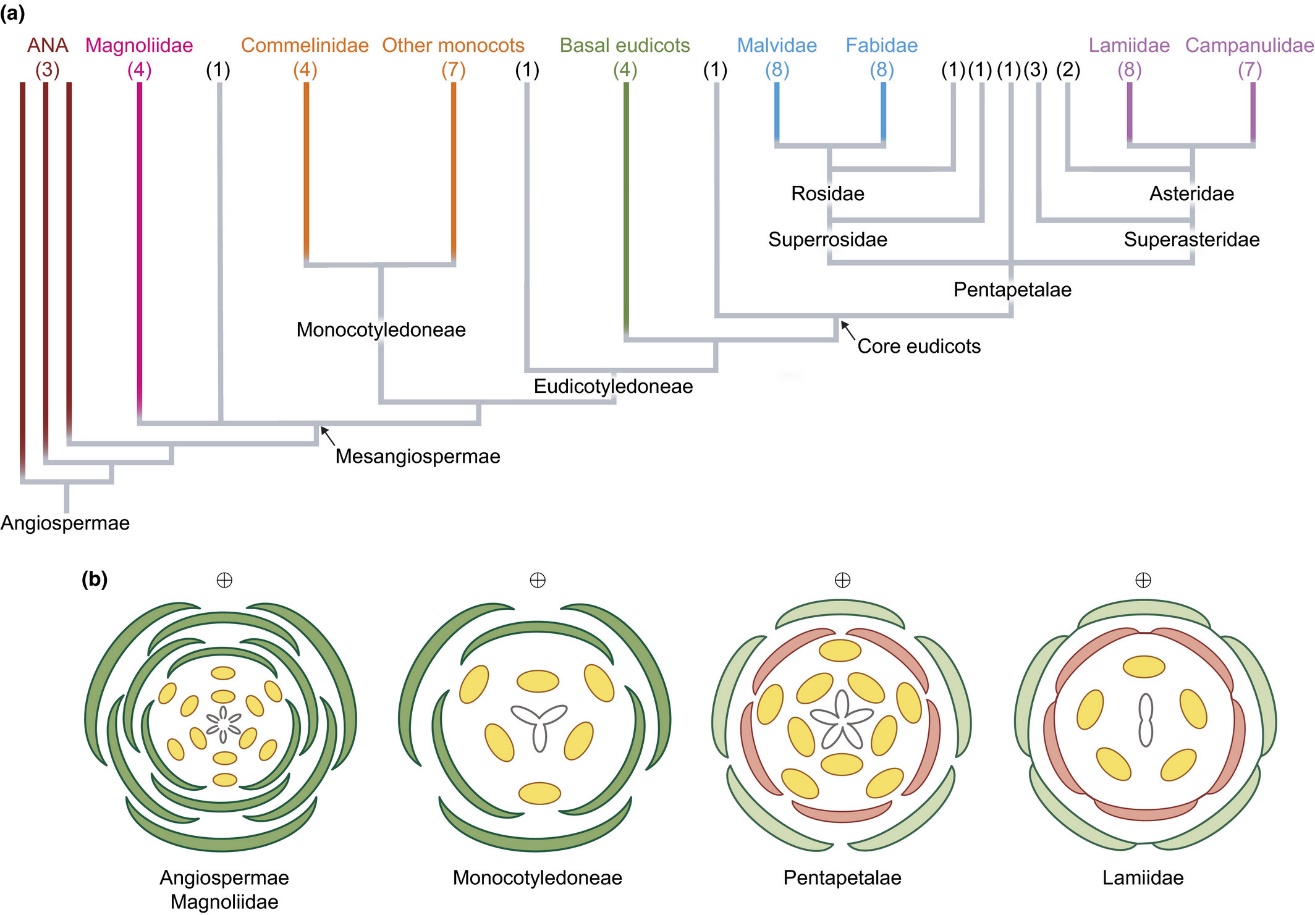
(a)簡化的被子植物生命之樹,分支頂部括號中的數字是該分支包含的“目”(order)的數量,注意將分支的名稱同下面(b)中的名稱對應起來;(b)中的示意圖被稱為“花圖示”,能很清楚地反映出花內各器官的排列方式:最中間的是雌蕊(心皮),黃色的圓點是雄蕊,雄蕊外面是花被片,如果花被片無法區分,則花萼和花瓣都畫成綠色,如果可以區分,那么內圈紅色的是花瓣,外圈綠色的是萼片(Smyth, 2018)
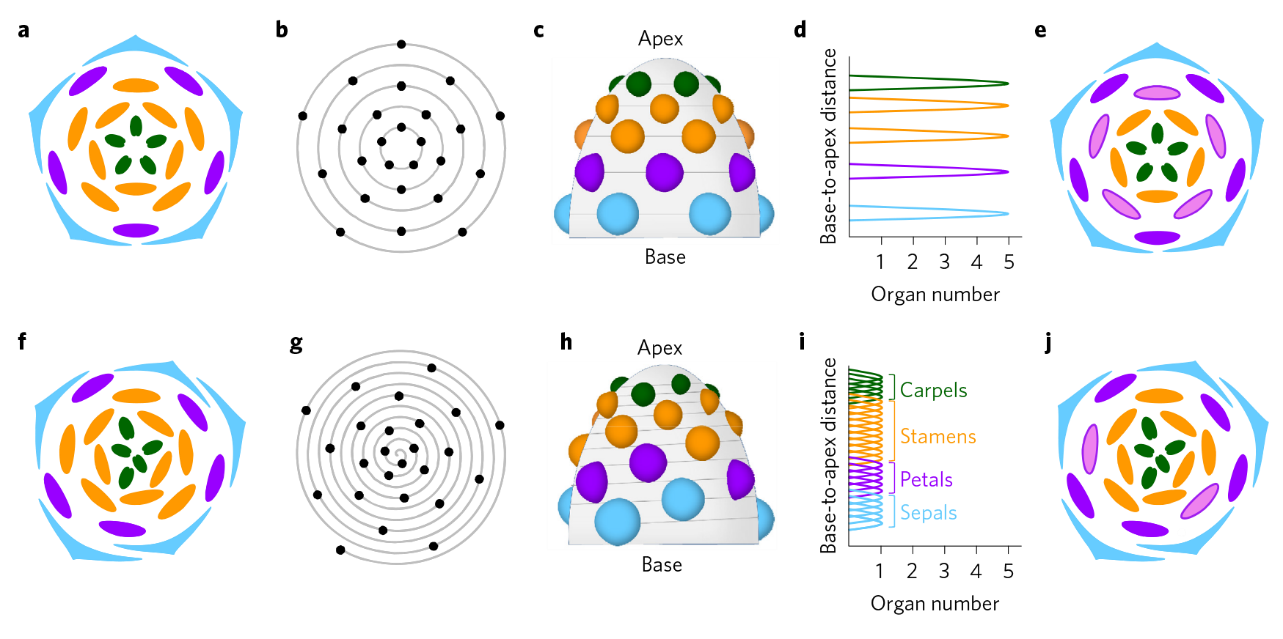
上排:輪狀花,下排:螺旋花;a和f:花圖示;c和h:花器官在花托上的空間分布示意圖;d和i:花器官數量沿花基部至頂端軸的變化;e和j:第三輪雄蕊瓣化時的花圖示(Wanget al., 2015)
(一)螺旋花
螺旋花的花瓣數量往往符合一個神奇的數學規律——斐波那契數列,又稱黃金分割數列,相鄰兩個花器官(如花瓣)之間形成137.5°的斐波那契角。據說是因為這樣排列葉片和花瓣不容易相互重疊,植物可以用更高的性價比獲得充足的陽光和花朵展示機會。
所謂斐波那契數列,就是從0、1開始,每個數字是前兩個數字的和,具體就是:0、1、1、2、3、5、8、13、21、34……由于這個數列開局是0和1,也就是一個偶數和一個奇數,偶數+奇數必然是奇數,所以第3個數一定是奇數;奇數(第2個數)+奇數(第3個數)=偶數(第4個數);奇數(第3個數)+偶數(第4個數)=奇數(第5個數)……總結一下,斐波那契數列的奇偶性排列一定是:偶奇奇偶奇奇偶奇奇……也就是“偶奇奇”循環。這還了得,奇數比偶數多!

螺旋著生的葉和花器官往往符合斐波那契數列。(A)云杉的螺旋葉序;(B)番茄頂芽;(C)非洲菊頭狀花序;(D)原基分化早期(Reinhardt & Gola, 2022)
然而這不意味著自然界中花瓣數量奇數的概率一定會更大,舉個例子:假設全世界只有四種花瓣數量類型——1、2、3、5,顯然,奇數比偶數多,但是花瓣數量=2的植物有幾萬種,其他花瓣數量的植物只有幾千種,那么哪怕“奇數陣營”多樣性高,它們在數量上成不了主流。因此需要考慮**“花瓣數量”這個性狀的數據分布**。這是個龐大的問題,小美不一定能在三天內查到所有數據,然而她可以采取化整為零的策略——螺旋花植物也沒多少,把有代表性的類群拉出來數數即可。

花器官發育順序。(a-c)輪生花,輪生花的花原基早起發育的時候也可能是螺旋開始的,可以連出斜列線(parastichies)和直列線(orthostichies);(d-f)螺旋花,只有斜列線沒有直列線,螺旋花和輪生花的發散角不同(Endress & Doyle, 2007)
花瓣螺旋著生的類群包括:五味子科(Schisandraceae)、苞被木科(Trimemiaceae)、蠟梅科(Calycanthaceae)、香皮茶科(Atherospermataceae)、毛茛科(Ranunculaceae)部分類群?、無油樟屬(Amborella)、八角屬(Illicium)、木蘭藤屬(Austrobaileya)、蓮屬(Nelumbo)、單心木蘭屬(Degeneria)、奇子樹屬(Idiospermum)、奎樂果屬(Gomortega)、八角桂屬(Hortonia)……
這都是些什么佶屈聱牙的名字?!其實這些類群中很多不分布在我國,如毛茛科中典型的螺旋花黑種草屬(Nigella)主要分布在歐洲,我國只有兩個引入觀賞的栽培種,而且黑種草屬看上去很像花瓣的部分其實是萼片,真正的花瓣藏在萼片和雄蕊之間,萼片和花瓣均以5瓣為多。
在這一堆螺旋花植物中,大部分人比較熟悉的有蠟梅和八角,如果解剖一下,會發現它們的花瓣數量確實不穩定(符合我們的要求),但是數起來有點費勁——苞片、萼片、花瓣之間有過渡狀態,不容易區分,不符合小美的計數規則。五味子科植物的花被片花瓣狀,5-24枚,數量也不確定,這似乎挺好,但是五味子還要上山去找,不太方便。
蓮花(Nelumbonucifera)是個不錯的選擇,不同品種的花瓣數量差異非常大,比如說“金秋”124瓣左右,“白花建蓮”16瓣左右,其雜交后代花瓣數量奇偶性無顯著差異。野生荷花花瓣數量在15-22之間,只要選對了蓮花品種或野生居群,不難roll出花瓣數量奇偶性均衡的數花瓣材料。
考慮到時間緊、預算少,先暫時把蓮花列入備選名單中。至于苞被木、無油樟之類就不考慮了,總不能為了數個花瓣數量申請標本傳遞吧?小帥何德何能?

白色分支主要是螺旋花被片、黑色分支主要是輪生花被片,其余介于兩者之間。輪生花被片不僅占據的分支多,分支內包含的物種總數更多(Endress & Doyle, 2007)
(二)輪生花
自然界中輪生花占大多數,其花瓣數量往往符合某種“花基數”,即一輪有n個花瓣,稱為n基數。常見的花基數包括:2、3、4、5、6,甚至7基數以及更高。一朵花可以有很多輪花瓣,最終花瓣數量就是n的倍數。顯然,輪狀花的花瓣數量由基數和輪數共同決定。常見的花基數和輪數是多少呢?

單子葉植物花主要為3基數,經常有兩輪,即6枚花瓣。雙子葉植物花主要為4、5基數,以5基數為多,很少有兩輪成10枚花瓣(野生的杜鵑花偶爾會突變,長出兩輪花瓣,形成10個花瓣裂片)。4基數可能是5基數花丟失一個基數導致的,2基數則有可能是3基數花丟失了一個基數導致的。6基數及以上的花比較少見。
常見植物,比如說蔥、百合、鳶尾、鴨跖草等等是3基數花,雙子葉植物中的木蘭科也是3基數花;桃、李、梅、櫻、杜鵑、海棠、薔薇、蜀葵、石竹等等是5基數花;桂花、油菜花、虞美人等等是4基數花。為什么3和5這兩個數字這么特別?有人認為可能是由于3和5剛好是斐波那契數列中的數字,如果輪狀花是從螺旋花進化來的,花基數傾向于斐波那契數列中的數字似乎說得過去,但是這個觀點尚未獲得證實。
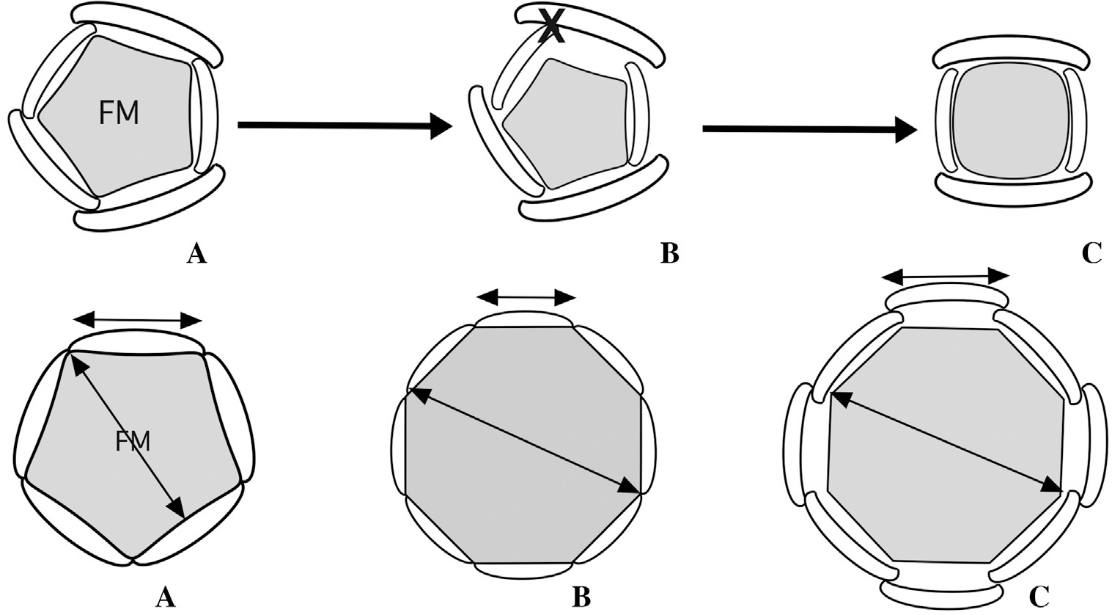
上排:(A)5基數花,(B)花分生組織縮小導致發育空間減少,丟失一個花器官,(C)4基數花;下排:(A)穩定的5基數花,(B)花分生組織增大、花器官減小,導致花基數增加而輪數不變,(C)花分生組織增大、花器官也增大,放不下這么多花器官,所以大家只能“錯身站著”,導致花器官重新排列,輪數增加,A和B都可能會產生C(De Craene, 2016)
總結來說,如果小美隨便在被子植物名錄里選一種植物來數花,那么她大概率得出的結果是奇數。瞧瞧!這還有公平性可言嗎?不如別數,直接接受小帥得了。別怕,我們還有Plan B。








技術路線B:栽培品種的不規則花瓣數量
同一種植物的花瓣數量并不是不變的,4瓣的植物偶爾也會長出5個花瓣,生長環境以及隨機等因素都有可能導致植物的花瓣數量發生波動,比如說低溫可以讓栽培的重瓣月季花長出更多花瓣、植物激素(如赤霉素和生長素)讓鳳仙花和香石竹花瓣增多等等。另外,花瓣越多、變數越多,對于螺旋花來說,如果你長了10圈花瓣,似乎再多長一兩圈也很正常;對于輪狀花來說,花基數越大,越不穩定。
人類在培育觀賞花卉品種時,會刻意選育一些花瓣多的品種,看上去雍容富貴。人類勢力的加入,給花瓣數量帶來了更多不確定性。為啥人工選育能弄出這么多花瓣?導致花瓣數量增加的原因很多,包括積累起源(主要針對離瓣花)、苞片起源(苞片或花萼變成類似花瓣的器官)、雌雄蕊起源(雌雄蕊瓣化)、臺閣起源(正常開放的花上層又開出一朵花),重復起源(花內層結構重復外層的結構和基數)和花序起源(針對單花組成的花序,如讓菊花外輪多圈小花變成舌狀小花)等,其中最重要的一種是:雌雄蕊瓣化。
雌雄蕊瓣化的底層原理比較復雜,這要從花瓣分化的本質,也就是基因調控說起——ABC模型,以及它的升級版ABCDE模型,其中研究最多的重瓣相關基因是C類和A類基因。簡單來說,就是某些基因突變后,在該長雄蕊的地方長出了花瓣、該長雌蕊的地方長出了花瓣,花瓣自然就變多了。染色體加倍等其他原因也可以產生重瓣花。另外,還有些基因突變可以改變花基數。
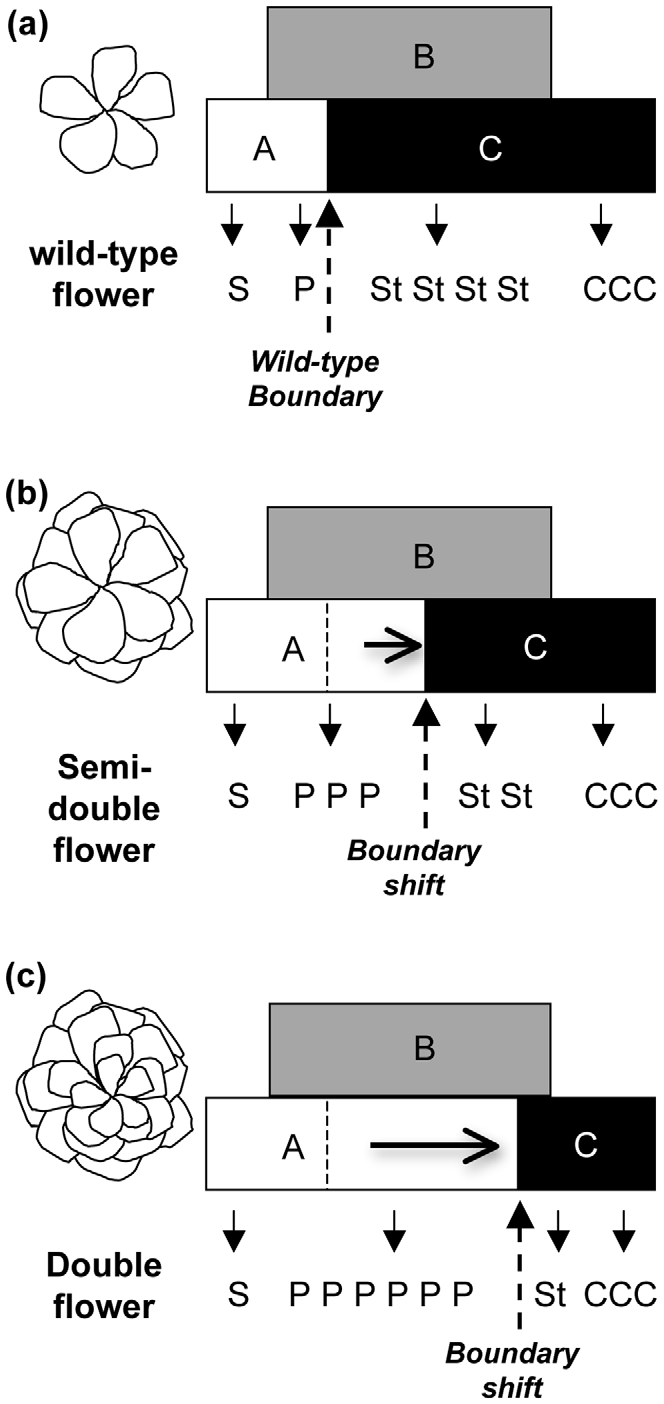
ABC模型對月季重瓣花形成的解釋。(a)野生型只有一輪花瓣;(b)半重瓣花和(c)重瓣花的花瓣和雄蕊邊界不穩定(Dubois et al. 2010)
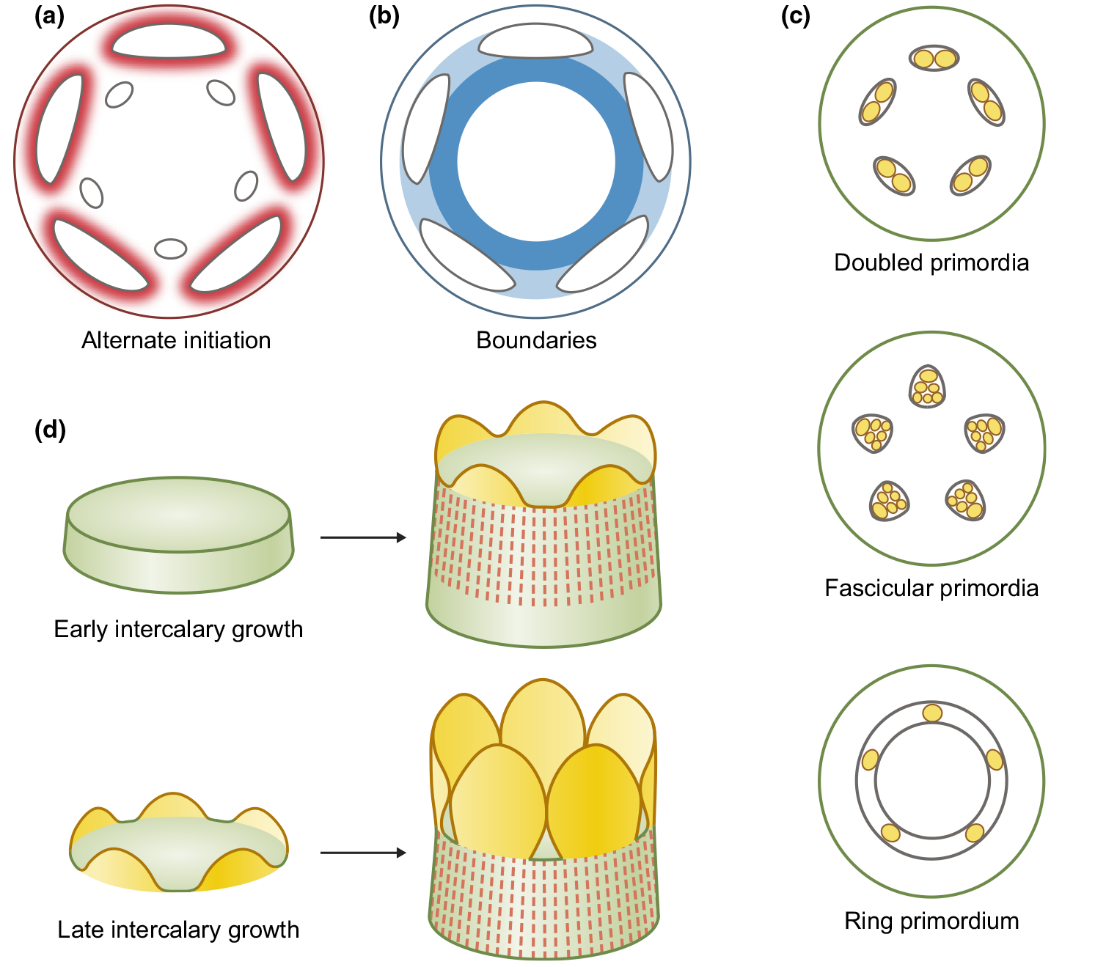
花器官排布的機制。(a)紅色表示生長素濃度,越高則越紅、對花器官分化的抑制作用越強,所以新的原基間隔著產生(白色的小橢圓形);(b)深藍色表示(a)圖中新原基的邊界;(c)雄蕊分化的時候經常在一個初始原基內部產生多個小原基(黃色的圓圈);(d)花器官基部融合的不同情況:花器官發育早期融合(上)和花器官發育一段時間后從原基下方發生融合(下)(Smyth, 2018)



小美又有話要說了,重瓣不就是加倍了嗎?無論是幾基數的花,兩輪就變成偶數了。一個品種,基數是確定的、重瓣的輪數是確定的,只要她選定了品種,那相當于直接選擇了花瓣數量的奇偶性,這還數什么?其實,重瓣有很多類型,不一定是整倍的,瓣化程度從低到高為:(1)單瓣→(2)半重瓣(花瓣層數少,保留較多正常雄蕊或雌蕊,仍然可育)→(3)牡丹型重瓣(結構松散,中心保留部分未完全瓣化的雄蕊)→(4)玫瑰型重瓣(雄蕊完全瓣化,不可育)。很少有研究會把重瓣花具體的花被片數量統計表公布出來,但是同一栽培環境下同一品種的花被片數量似乎是有規律的,比如說東方百合的3個品種最大出現頻率的正常花被片數量分別為:“Siberia”最多為5或6片、“Sorbonne”最多為5片、“Fastrada”最多為6片。
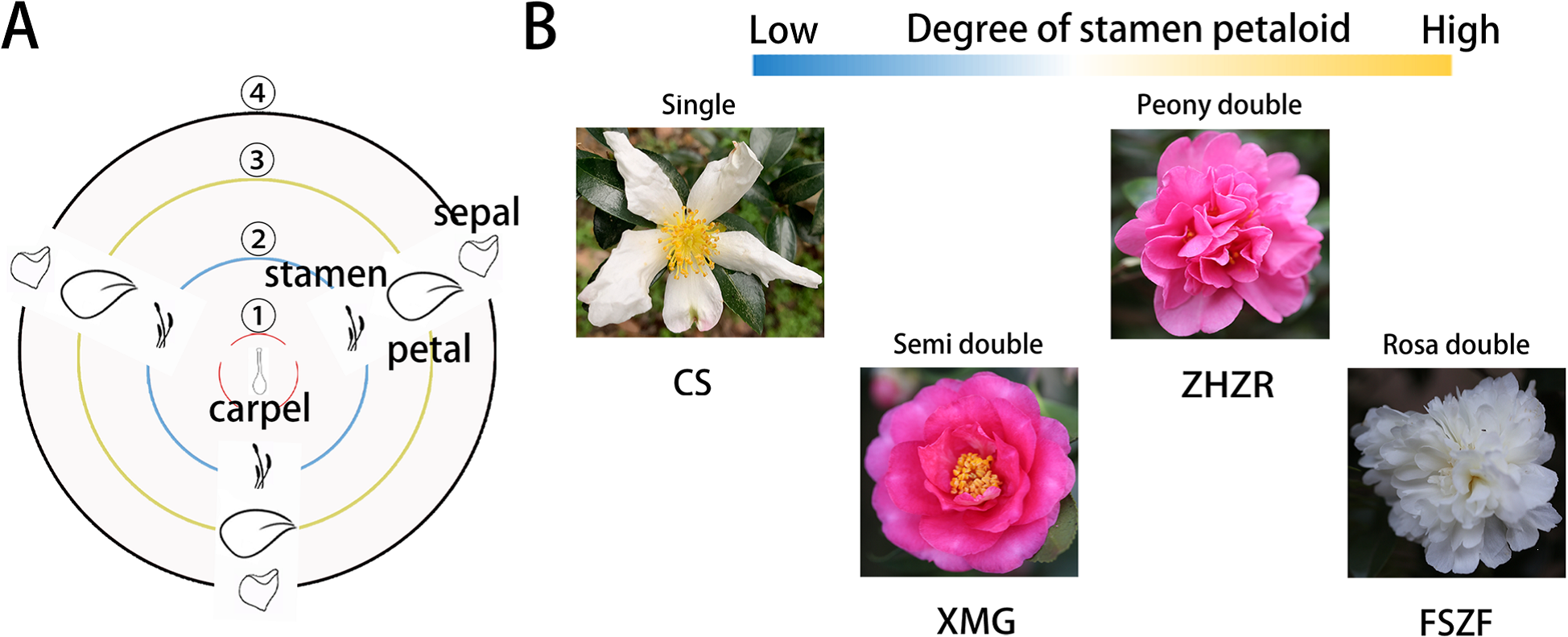
山茶花單瓣至重瓣的瓣化程度(Fan et al., 2022)



啊!天哪!數個花瓣怎么這么復雜!我們已經就這個問題討論4千多字,快趕上一篇小論文的字數了!不如隨便選一種花瓣奇偶不確定的花,先買來統計下,如果奇偶數分布是對稱的(即奇數和偶數概率相同),那我們就正式用這種花的花瓣數量來決定是否去約會,如果奇偶數不對稱就換一個品種,直到滿足我們的要求。
栽培的重瓣月季花/玫瑰花就是很好的選擇!同一個品種“月月粉”有的研究中數出來是23片花瓣,有的研究數出來是20片花瓣。當然,如果小美有閑情也可以去大觀公園趴欄桿上數幾天荷花,顯然她沒什么閑情。
現在我們可以敲定最終方案:1)網購35朵月季花(玫瑰花);2)用其中30朵統計出花瓣數量的奇偶性分布,如果花瓣數量奇、偶數出現的概率顯著偏離0.5,就重新購買一批月季花,直到該批次奇偶性呈現出比較均勻的分布;3)使用剩下的5朵花統計花朵數量奇偶性,如超過3朵為奇數,就選擇接受小帥的邀請,反之就拒絕。
小美舒了口氣:不愧是我,簡直滴水不漏,希望小帥可以耐心等待一批奇偶均勻的月季花到貨。
果然,知識改變命運。
【完】
感謝讀到最后,這是“室花小美的脫單之路”系列的第一篇(不保證有第二篇)。植物的花瓣數量是一個有趣的研究方向,有各種不同的研究角度,比如說:調控基因、生理代謝過程、花原基發育、花瓣數量在整個植物進化歷史中的系統發生過程等等,感興趣的讀者可以去搜索一下文獻,某些知識分享網站上也有一些非常專業的回答。
下一篇再見!
參考文獻:
蔡水花, 馬文寶, 周曉, 等. 野生寶興杜鵑重瓣花的發現及其形成機制探討[J]. 四川林業科技, 2022, 43(01): 121-126.
付彥榮. 中國北方野生蓮(Nelumbo nucifera)種質多樣性研究[D]. 北京林業大學, 2021.
劉引. 月季種質資源評價及部分觀賞性狀QTL定位研究[D]. 河南農業大學, 2021.
彭潔, 劉引, 武榮花, 等. 重瓣花及其分子機制的研究進展[J]. 中國農學通報, 2023, 39(19): 65-72.
戚雨岑, 楊淑敏, 蘇琳, 等. RcAGL61基因調控雄蕊和花瓣之間轉變影響月季花瓣數量[J]. 西北植物學報, 2024, 44(06): 904-911.
孫明偉, 邵小斌, 趙統利, 等. 東方百合花器官數量變異分析[J]. 福建農業學報, 2018, 33(04): 373-380.
王宏哲, 張睿, 程劼, 等. 花基本結構的多樣性及其分子機制[J]. 中國科學: 生命科學, 2019,49(04): 292-300.
王其超, 陳耀東, 辛春德, 等. 黑龍江野生荷花資源考察[J]. 中國園林, 1997(04): 39-41.
嚴寒松. 中國蓮高密度遺傳圖譜的構建及花瓣數控制基因的定位[D]. 福建農林大學, 2019.
趙印泉, 劉青林. 重瓣花的形成機理及遺傳特性研究進展[J]. 西北植物學報, 2009, 29(04): 832-841.
Annick D, OlivierR, Marion M, et al. Tinkering with the C-function: a molecular frame for theselection of double flowers in cultivated roses [J]. PloS one, 2010, 5(2): e9288.
De Craene LR. Meristicchanges in flowering plants: How flowers play with numbers[J]. Flora, 2016, 22-37.
Endress PK. TheFlowers in Extant Basal Angiosperms and Inferences on Ancestral Flowers[J]. InternationalJournal of Plant Sciences, 2001, 162(5): 1111-1140.
Endress PK,Doyle JA. Floral phyllotaxis in basal angiosperms: development andevolution[J]. Current opinion in plant biology, 2007, 10(1): 52-7.
Fan ML, Li XL,Zhang J, et al. Floral organ transcriptome in Camellia sasanqua providedinsight into stamen petaloid[J]. BMC Plant Biology, 2022, 22(1): 474-474.
Nan M, Wen C, TiangangF, et al. Low temperature-induced DNA hypermethylation attenuates expression ofRhAG, an AGAMOUS homolog, and increases petal number in rose (Rosa hybrida)[J]. BMC plant biology, 2015, 15(1): 237.
Reinhardt D,Gola EM. Law and order in plants - the origin and functional relevance ofphyllotaxis[J]. Trends in plant science, 2022, 27(10): 1017-1032.
Rümpler F, Theissen G. Reconstructing the ancestral flower of extantangiosperms: the 'war of the whorls' is heating up[J]. Journal of experimentalbotany, 2019, 70(10): 2615-2622.
Smyth DR. Evolutionand genetic control of the floral ground plan[J]. The New phytologist, 2018, 220(1):70-86.
Wang PP, Liao H, Zhang WG, et al. Flexibility inthe structure of spiral flowers and its underlying mechanisms[J]. NaturePlants, 2016, 2(1): 15188.
來源: 昆明植物園
